

Conserved Ser/Arg-rich motif in PPZ orthologs from fungi is important for its role in cation tolerance
- PMID: 22232558
- PMCID: PMC3293525
- DOI: 10.1074/jbc.M111.299438
Conserved Ser/Arg-rich motif in PPZ orthologs from fungi is important for its role in cation tolerance
Abstract
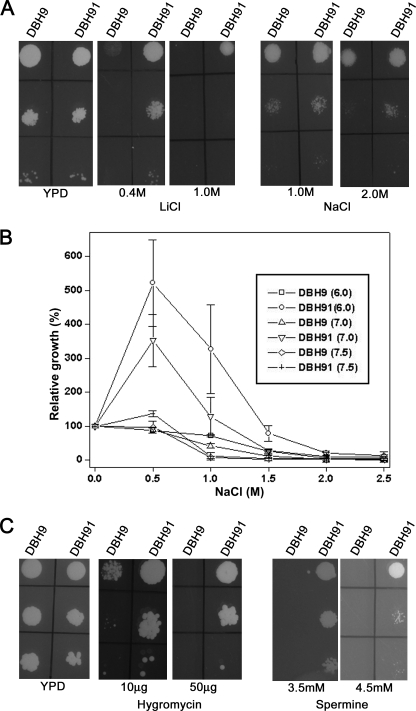
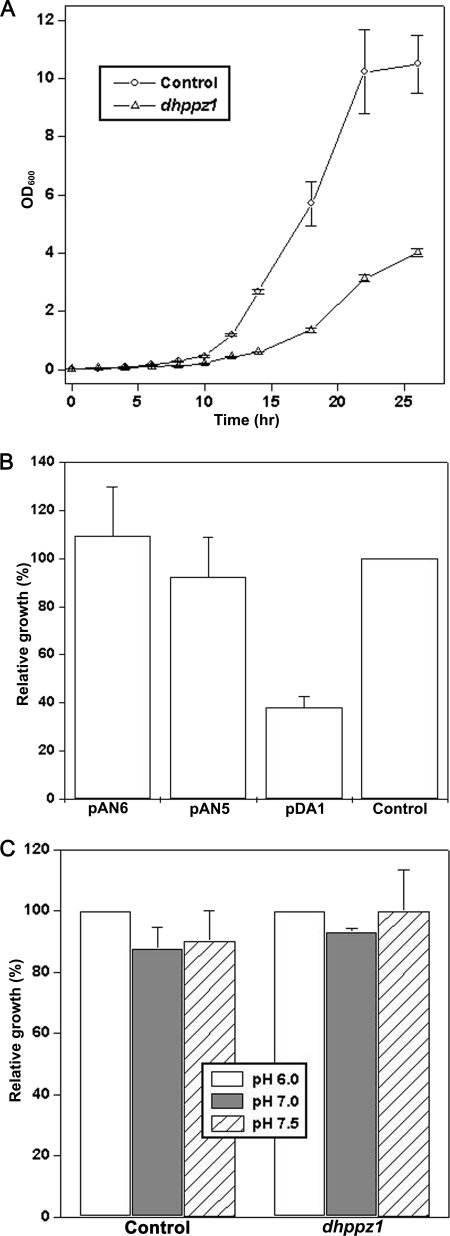
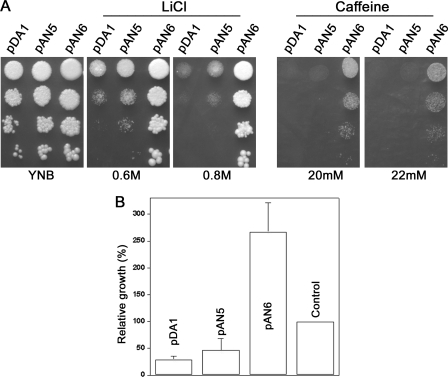
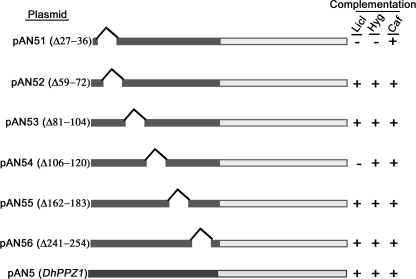
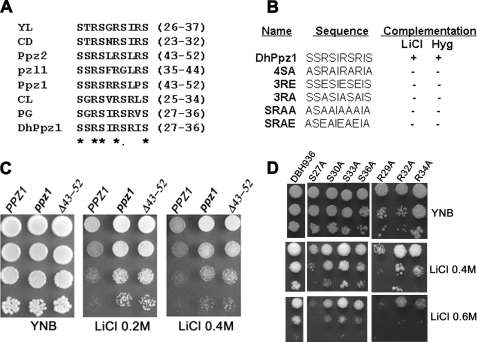
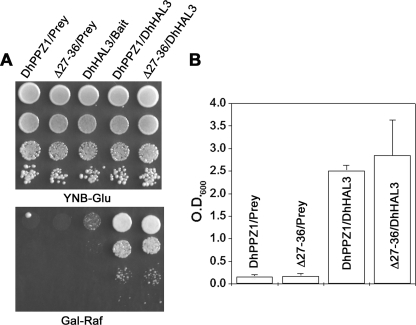
PPZ1 orthologs, novel members of a phosphoprotein phosphatase family of phosphatases, are found only in fungi. They regulate diverse physiological processes in fungi e.g. ion homeostasis, cell size, cell integrity, etc. Although they are an important determinant of salt tolerance in fungi, their physiological role remained unexplored in any halotolerant species. In this context we report here molecular and functional characterization of DhPPZ1 from Debaryomyces hansenii, which is one of the most halotolerant and osmotolerant species of yeast. Our results showed that DhPPZ1 knock-out strain displayed higher tolerance to toxic cations, and unlike in Saccharomyces cerevisiae, Na(+)/H(+) antiporter appeared to have an important role in this process. Besides salt tolerance, DhPPZ1 also had role in cell wall integrity and growth in D. hansenii. We have also identified a short, serine-arginine-rich sequence motif in DhPpz1p that is essential for its role in salt tolerance but not in other physiological processes. Taken together, these results underscore a distinct role of DhPpz1p in D. hansenii and illustrate an example of how organisms utilize the same molecular tool box differently to garner adaptive fitness for their respective ecological niches.
Figures

References
-
- Cohen P. T. (1997) Novel protein serine/threonine phosphatases. Variety is the spice of life. Trends Biochem. Sci. 22, 245–251 - PubMed
-
- Cohen P. T. (2004) Protein Phosphatases (Arino J., ed.) pp. 1–20, Springer, Berlin
-
- Moorhead G. B., De Wever V., Templeton G., Kerk D. (2009) Evolution of protein phosphatases in plants and animals. Biochem. J. 417, 401–409 - PubMed
-
- Shi Y. (2009) Serine/threonine phosphatases. Mechanism through structure. Cell 139, 468–484 - PubMed
-
- Blatch G. L., Lässle M. (1999) The tetratricopeptide repeat. A structural motif mediating protein-protein interactions. Bioessays 21, 932–939 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
