

Dynein-dependent processive chromosome motions promote homologous pairing in C. elegans meiosis
- PMID: 22232701
- PMCID: PMC3255982
- DOI: 10.1083/jcb.201106022
Dynein-dependent processive chromosome motions promote homologous pairing in C. elegans meiosis
Abstract
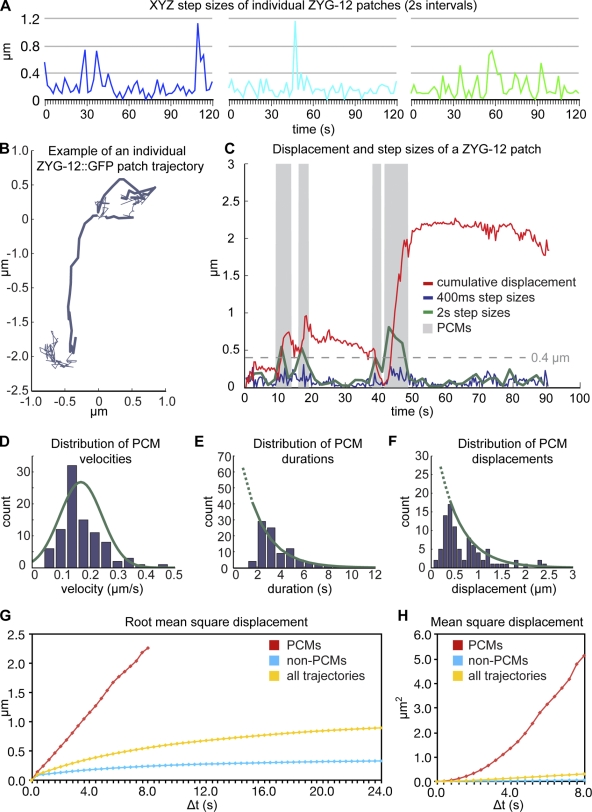
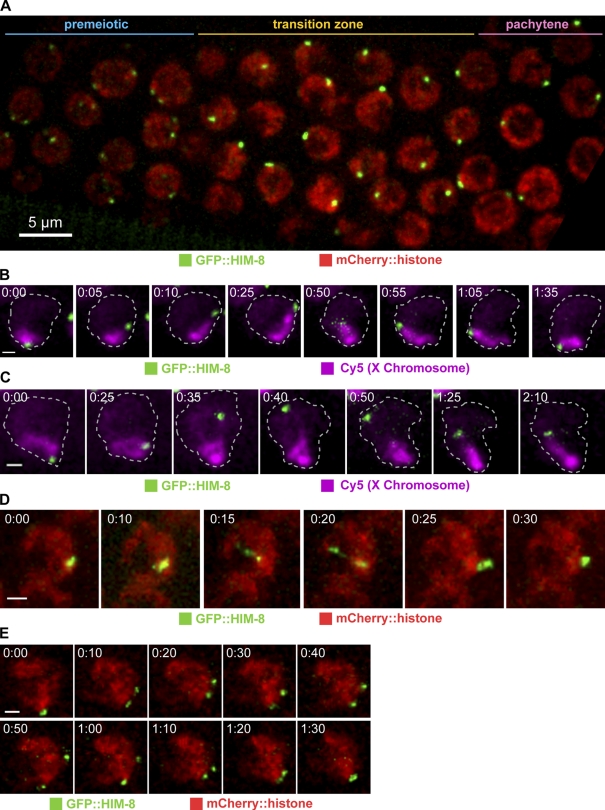
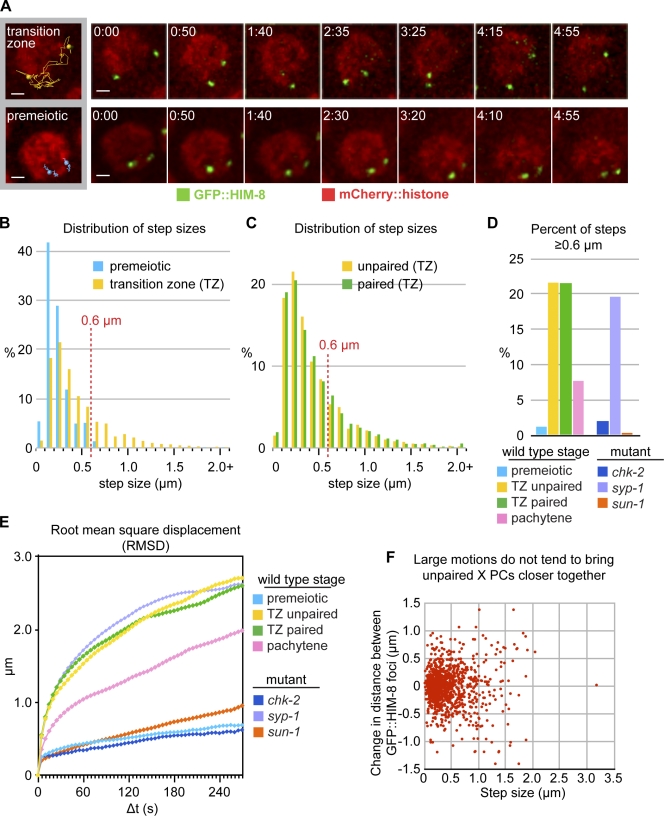
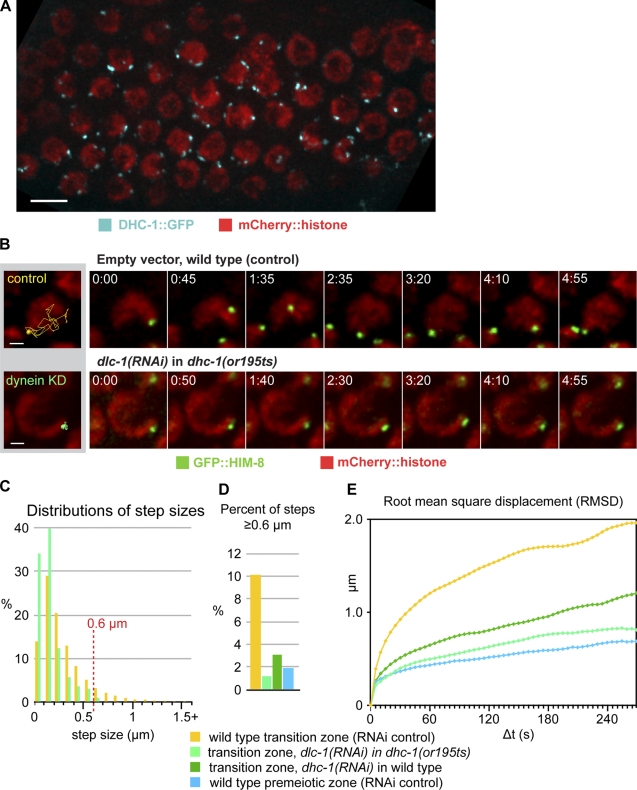
Meiotic chromosome segregation requires homologue pairing, synapsis, and crossover recombination, which occur during meiotic prophase. Telomere-led chromosome motion has been observed or inferred to occur during this stage in diverse species, but its mechanism and function remain enigmatic. In Caenorhabditis elegans, special chromosome regions known as pairing centers (PCs), rather than telomeres, associate with the nuclear envelope (NE) and the microtubule cytoskeleton. In this paper, we investigate chromosome dynamics in living animals through high-resolution four-dimensional fluorescence imaging and quantitative motion analysis. We find that chromosome movement is constrained before meiosis. Upon prophase onset, constraints are relaxed, and PCs initiate saltatory, processive, dynein-dependent motions along the NE. These dramatic motions are dispensable for homologous pairing and continue until synapsis is completed. These observations are consistent with the idea that motions facilitate pairing by enhancing the search rate but that their primary function is to trigger synapsis. This quantitative analysis of chromosome dynamics in a living animal extends our understanding of the mechanisms governing faithful genome inheritance.
Figures



References
-
- Bass H.W., Riera-Lizarazu O., Ananiev E.V., Bordoli S.J., Rines H.W., Phillips R.L., Sedat J.W., Agard D.A., Cande W.Z. 2000. Evidence for the coincident initiation of homolog pairing and synapsis during the telomere-clustering (bouquet) stage of meiotic prophase. J. Cell Sci. 113:1033–1042 - PubMed
-
- Baudrimont A., Penkner A., Woglar A., Machacek T., Wegrostek C., Gloggnitzer J., Fridkin A., Klein F., Gruenbaum Y., Pasierbek P., Jantsch V. 2010. Leptotene/zygotene chromosome movement via the SUN/KASH protein bridge in Caenorhabditis elegans. PLoS Genet. 6:e1001219 10.1371/journal.pgen.1001219 - DOI - PMC - PubMed
-
- Carlton P.M., Boulanger J., Kervrann C., Sibarita J.B., Salamero J., Gordon-Messer S., Bressan D., Haber J.E., Haase S., Shao L., et al. 2010. Fast live simultaneous multiwavelength four-dimensional optical microscopy. Proc. Natl. Acad. Sci. USA. 107:16016–16022 10.1073/pnas.1004037107 - DOI - PMC - PubMed
-
- Chen, H., W.K. Clyborne, J.W. Sedat, and D.A. Agard. 1992. PRIISM: an integrated system for display and analysis of 3-D microscope images. In Biomedical Image Processing and Three-Dimensional Microscopy. Vol. 784. R.S. Acharya, C.J. Cogswell, and D.B. Goldgof, editors. SPIE, Bellingham, WA. 10.1117/12.59604.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
