

The left occipitotemporal cortex does not show preferential activity for words
- PMID: 22235035
- PMCID: PMC3491762
- DOI: 10.1093/cercor/bhr295
The left occipitotemporal cortex does not show preferential activity for words
Abstract
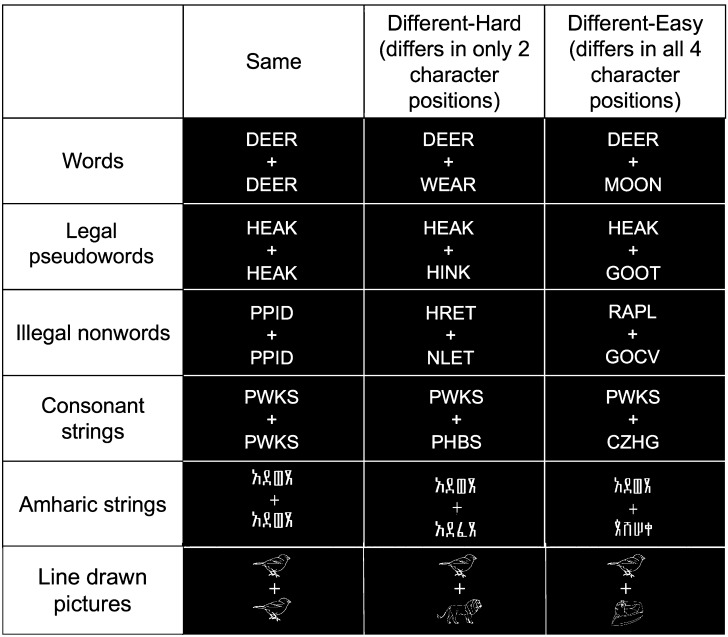
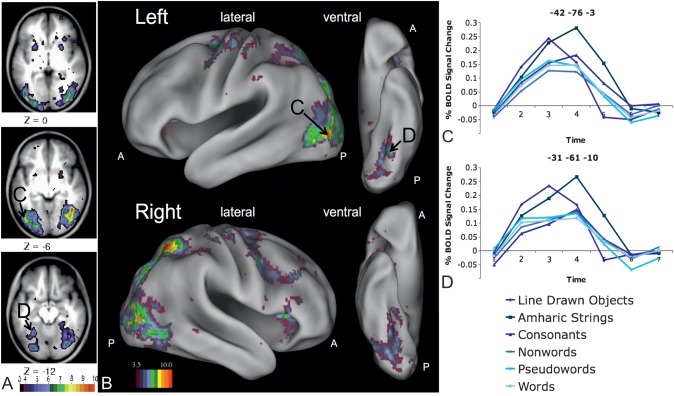
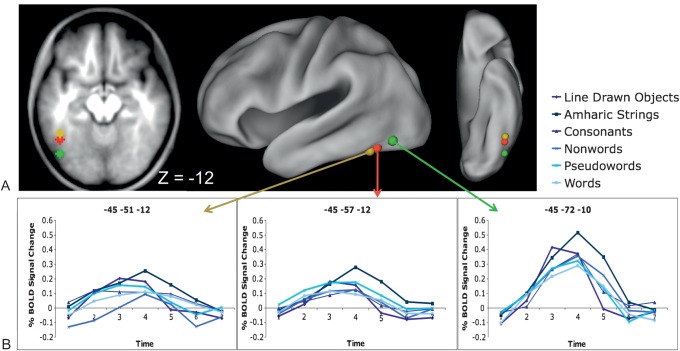
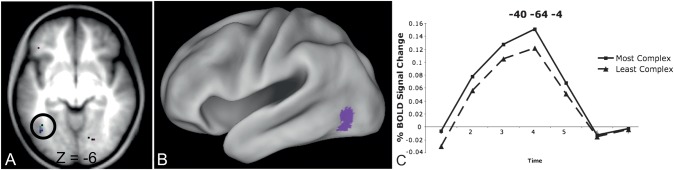
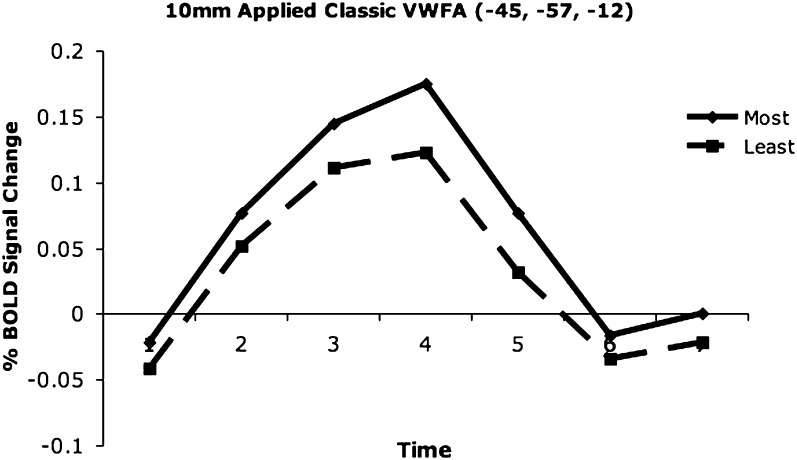
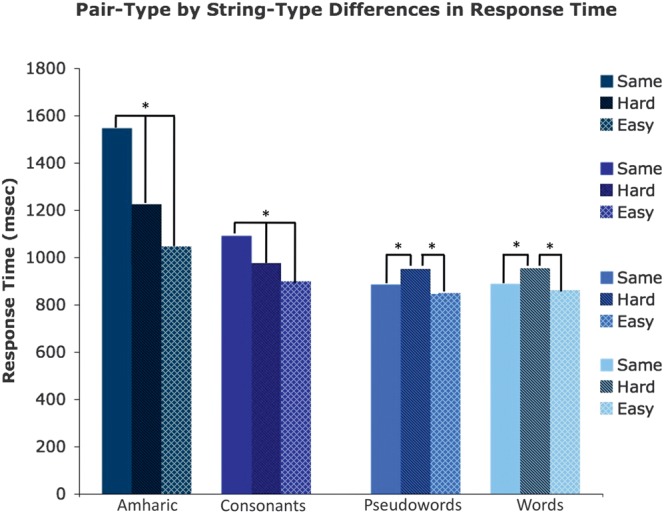
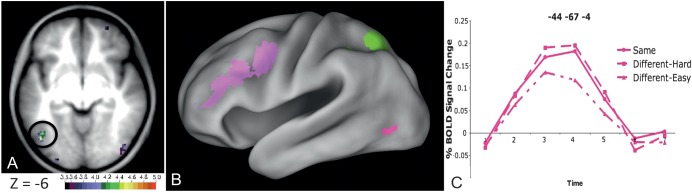
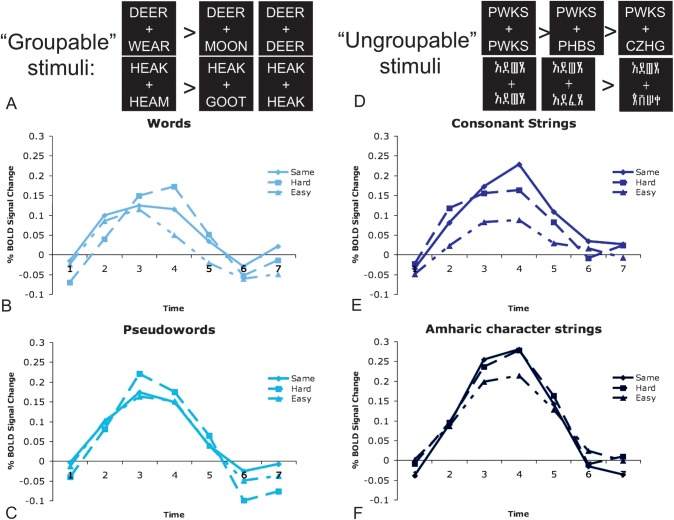
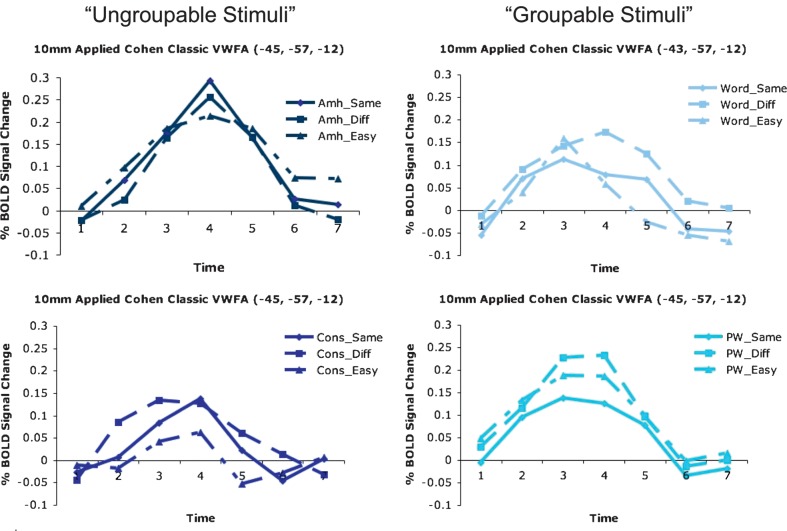
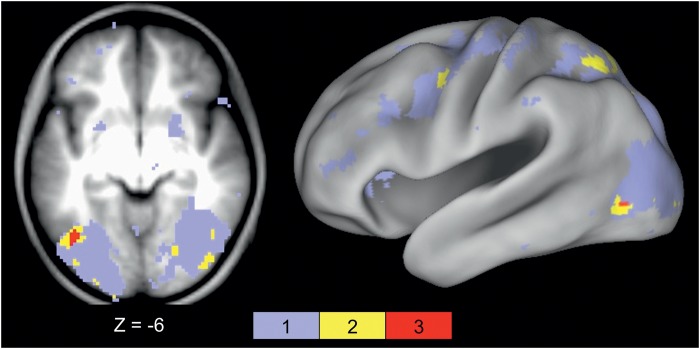
Regions in left occipitotemporal (OT) cortex, including the putative visual word form area, are among the most commonly activated in imaging studies of single-word reading. It remains unclear whether this part of the brain is more precisely characterized as specialized for words and/or letters or contains more general-use visual regions having properties useful for processing word stimuli, among others. In Analysis 1, we found no evidence of greater activity in left OT regions for words or letter strings relative to other high-spatial frequency high-contrast stimuli, including line drawings and Amharic strings (which constitute the Ethiopian writing system). In Analysis 2, we further investigated processing characteristics of OT cortex potentially useful in reading. Analysis 2 showed that a specific part of OT cortex 1) is responsive to visual feature complexity, measured by the number of strokes forming groups of letters or Amharic strings and 2) processes learned combinations of characters, such as those in words and pseudowords, as groups but does not do so in consonant and Amharic strings. Together, these results indicate that while regions of left OT cortex are not specialized for words, at least part of OT cortex has properties particularly useful for processing words and letters.
Figures
References
-
- Bar M, Tootell RB, Schacter DL, Greve DN, Fischl B, Mendola JD, Rosen BR, Dale AM. Cortical mechanisms specific to explicit visual object recognition. Neuron. 2001;29:529–535. - PubMed
-
- Behrmann M, Black SE, Bub D. The evolution of pure alexia: a longitudinal study of recovery. Brain Lang. 1990;39:405–427. - PubMed
-
- Behrmann M, Nelson J, Sekuler EB. Visual complexity in letter-by-letter reading: “pure” alexia is not pure. Neuropsychologia. 1998;36:1115–1132. - PubMed
-
- Ben-Shachar M, Dougherty RF, Deutsch GK, Wandell BA. Differential sensitivity to words and shapes in ventral occipito-temporal cortex. Cereb Cortex. 2007;17:1604–1611. - PubMed
