

Knockdown of ZNF268, which is transcriptionally downregulated by GATA-1, promotes proliferation of K562 cells
- PMID: 22235304
- PMCID: PMC3250450
- DOI: 10.1371/journal.pone.0029518
Knockdown of ZNF268, which is transcriptionally downregulated by GATA-1, promotes proliferation of K562 cells
Abstract
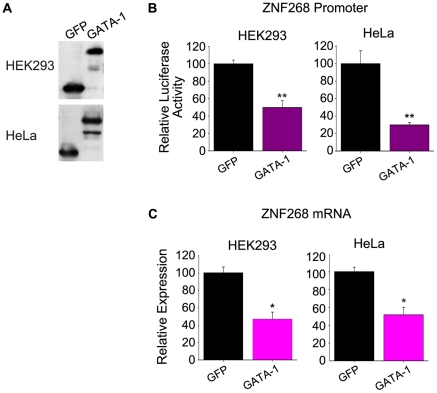
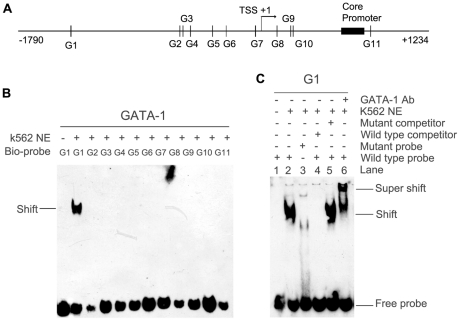
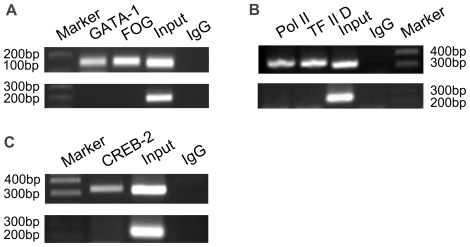
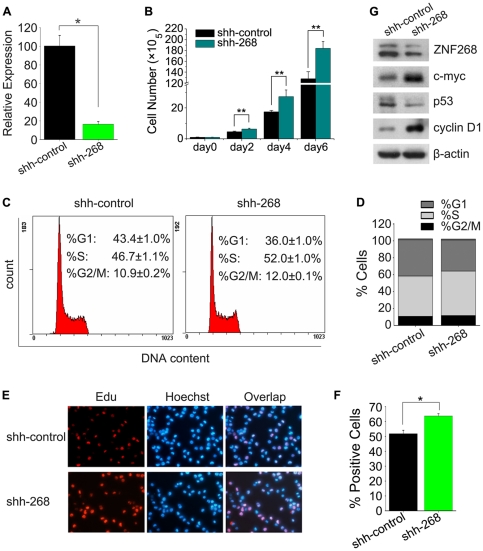
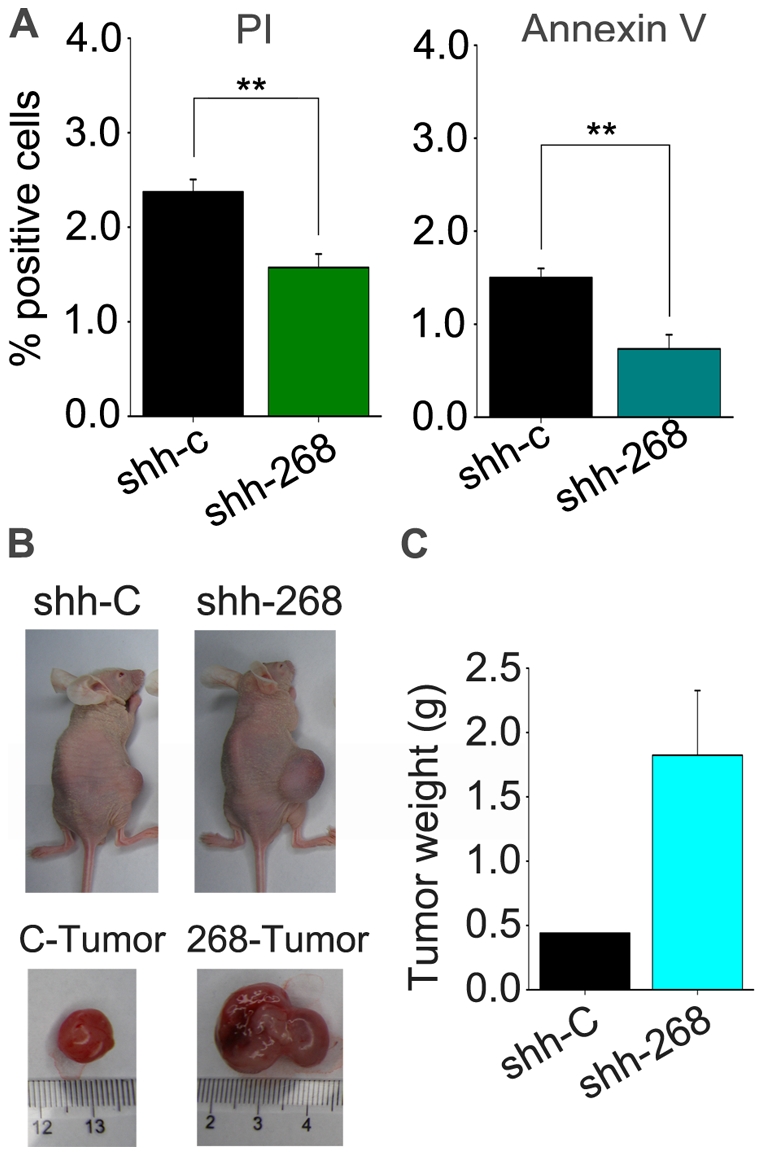
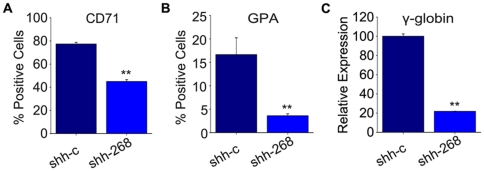
The human ZNF268 gene encodes a typical KRAB-C2H2 zinc finger protein that may participate in hematopoiesis and leukemogenesis. A recent microarray study revealed that ZNF268 expression continuously decreases during erythropoiesis. However, the molecular mechanisms underlying regulation of ZNF268 during hematopoiesis are not well understood. Here we found that GATA-1, a master regulator of erythropoiesis, repressed the promoter activity and transcription of ZNF268. Electrophoretic mobility shift assays and chromatin immunoprecipitation assays showed that GATA-1 directly bound to a GATA binding site in the ZNF268 promoter in vitro and in vivo. Knockdown of ZNF268 in K562 erythroleukemia cells with specific siRNA accelerated cellular proliferation, suppressed apoptosis, and reduced expression of erythroid-specific developmental markers. It also promoted growth of subcutaneous K562-derived tumors in nude mice. These results suggest that ZNF268 is a crucial downstream target and effector of GATA-1. They also suggest the downregulation of ZNF268 by GATA-1 is important in promoting the growth and suppressing the differentiation of K562 erythroleukemia cells.
Conflict of interest statement
Figures
Similar articles
-
GATA-1 directly regulates p21 gene expression during erythroid differentiation.Cell Cycle. 2010 May 15;9(10):1972-80. doi: 10.4161/cc.9.10.11602. Epub 2010 May 15. Cell Cycle. 2010. PMID: 20495378 Free PMC article.
-
Role of transcriptional corepressor ETO2 in erythroid cells.Exp Hematol. 2013 Mar;41(3):303-15.e1. doi: 10.1016/j.exphem.2012.10.015. Epub 2012 Nov 2. Exp Hematol. 2013. PMID: 23127762
-
Transcription of human zinc finger ZNF268 gene requires an intragenic promoter element.J Biol Chem. 2006 Aug 25;281(34):24623-36. doi: 10.1074/jbc.M602753200. Epub 2006 Jun 20. J Biol Chem. 2006. PMID: 16787922
-
Role of STAT3 and GATA-1 interactions in gamma-globin gene expression.Exp Hematol. 2009 Aug;37(8):889-900. doi: 10.1016/j.exphem.2009.05.004. Epub 2009 May 15. Exp Hematol. 2009. PMID: 19447160 Free PMC article.
-
Isolation and characterization of hematopoietic transcription factor complexes by in vivo biotinylation tagging and mass spectrometry.Ann N Y Acad Sci. 2005;1054:55-67. doi: 10.1196/annals.1345.008. Ann N Y Acad Sci. 2005. PMID: 16339652 Review.
Cited by
-
Myocardial ischemic preconditioning upregulated protein 1(Mipu1):zinc finger protein 667 - a multifunctional KRAB/C2H2 zinc finger protein.Braz J Med Biol Res. 2015 Jan;48(1):1-5. doi: 10.1590/1414-431x20144029. Epub 2014 Oct 31. Braz J Med Biol Res. 2015. PMID: 25493376 Free PMC article.
-
An expanded role for dipeptidyl peptidase 4 in cell regulation.Curr Opin Hematol. 2020 Jul;27(4):215-224. doi: 10.1097/MOH.0000000000000590. Curr Opin Hematol. 2020. PMID: 32487805 Free PMC article. Review.
-
ZNF300 knockdown inhibits forced megakaryocytic differentiation by phorbol and erythrocytic differentiation by arabinofuranosyl cytidine in K562 cells.PLoS One. 2014 Dec 8;9(12):e114768. doi: 10.1371/journal.pone.0114768. eCollection 2014. PLoS One. 2014. PMID: 25485965 Free PMC article.
-
hERG potassium channel blockage by scorpion toxin BmKKx2 enhances erythroid differentiation of human leukemia cells K562.PLoS One. 2013 Dec 26;8(12):e84903. doi: 10.1371/journal.pone.0084903. eCollection 2013. PLoS One. 2013. PMID: 24386436 Free PMC article.
-
The zinc finger protein ZNF268 is overexpressed in human cervical cancer and contributes to tumorigenesis via enhancing NF-κB signaling.J Biol Chem. 2012 Dec 14;287(51):42856-66. doi: 10.1074/jbc.M112.399923. Epub 2012 Oct 22. J Biol Chem. 2012. PMID: 23091055 Free PMC article.
References
-
- Gou DM, Sun Y, Gao L, Chow LM, Huang J, et al. Cloning and characterization of a novel Kruppel-like zinc finger gene, ZNF268, expressed in early human embryo. Biochim Biophys Acta. 2001;1518:306–310. - PubMed
-
- Sun C, Zhao ZZ, Gao L, Sun Y, Shao HJ, et al. [Cloning and characterization of two novel alternatively spliced transcripts of ZNF268]. Yi Chuan. 2006;28:513–517. - PubMed
-
- Shao H, Zhu C, Zhao Z, Guo M, Qiu H, et al. KRAB-containing zinc finger gene ZNF268 encodes multiple alternatively spliced isoforms that contain transcription regulatory domains. Int J Mol Med. 2006;18:457–463. - PubMed
-
- Sun Y, Shao H, Li Z, Liu J, Gao L, et al. ZNF268, a novel kruppel-like zinc finger protein, is implicated in early human liver development. Int J Mol Med. 2004;14:971–975. - PubMed
-
- Krackhardt AM, Witzens M, Harig S, Hodi FS, Zauls AJ, et al. Identification of tumor-associated antigens in chronic lymphocytic leukemia by SEREX. Blood. 2002;100:2123–2131. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
