

Arabidopsis plants grown in the field and climate chambers significantly differ in leaf morphology and photosystem components
- PMID: 22236032
- PMCID: PMC3296669
- DOI: 10.1186/1471-2229-12-6
Arabidopsis plants grown in the field and climate chambers significantly differ in leaf morphology and photosystem components
Abstract
Background: Plants exhibit phenotypic plasticity and respond to differences in environmental conditions by acclimation. We have systematically compared leaves of Arabidopsis thaliana plants grown in the field and under controlled low, normal and high light conditions in the laboratory to determine their most prominent phenotypic differences.
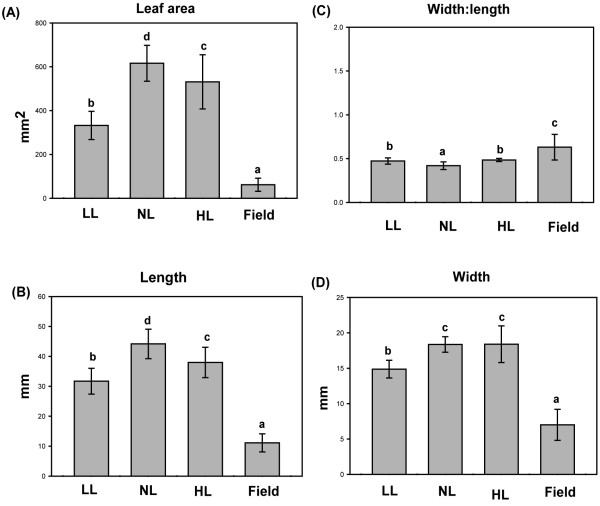
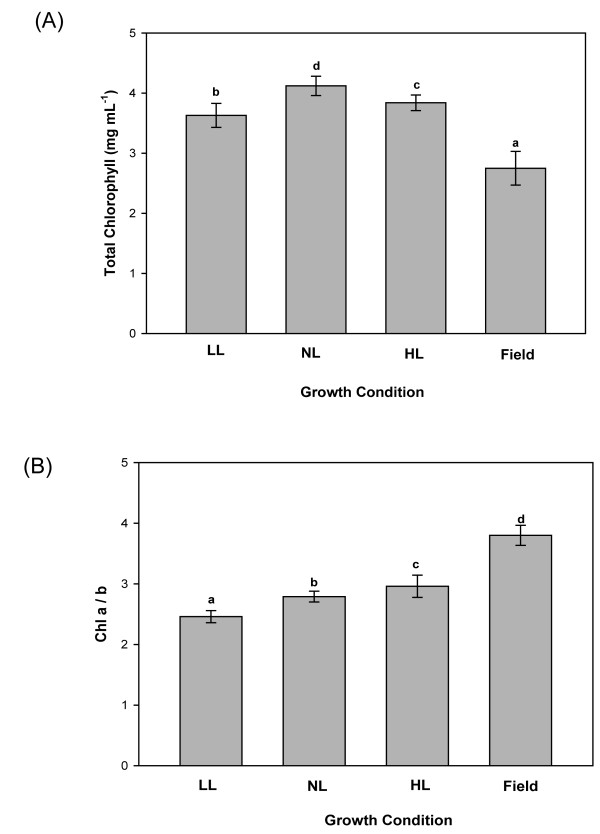
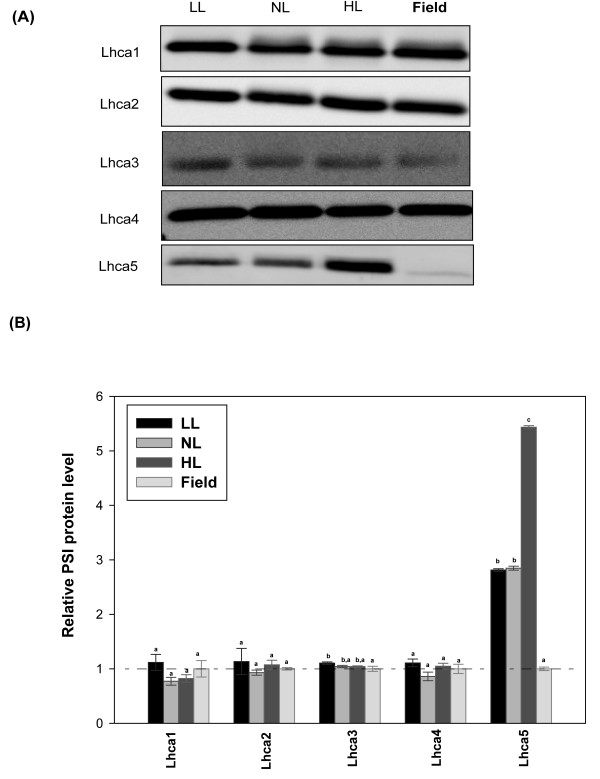
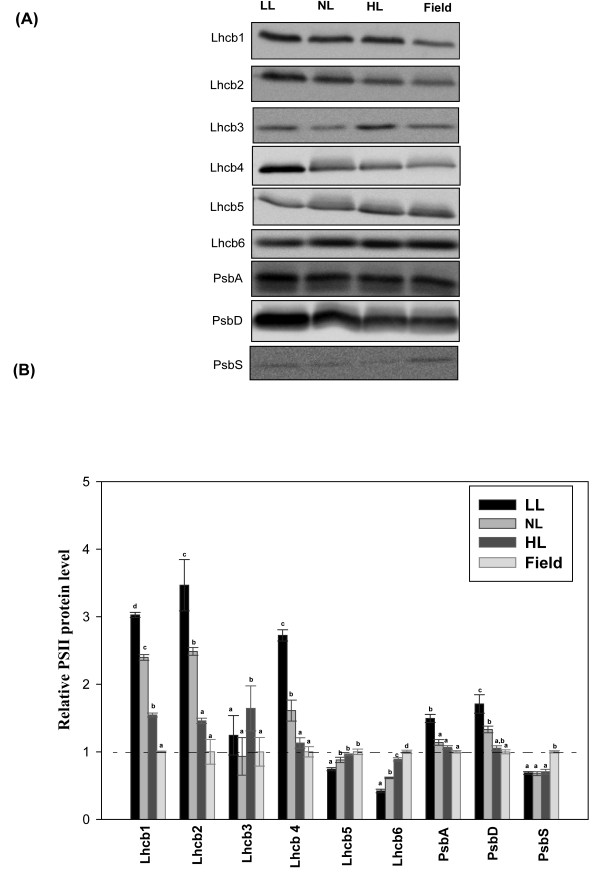
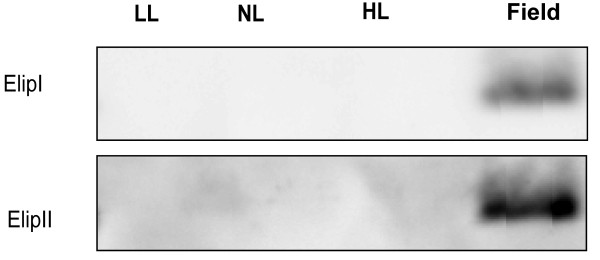
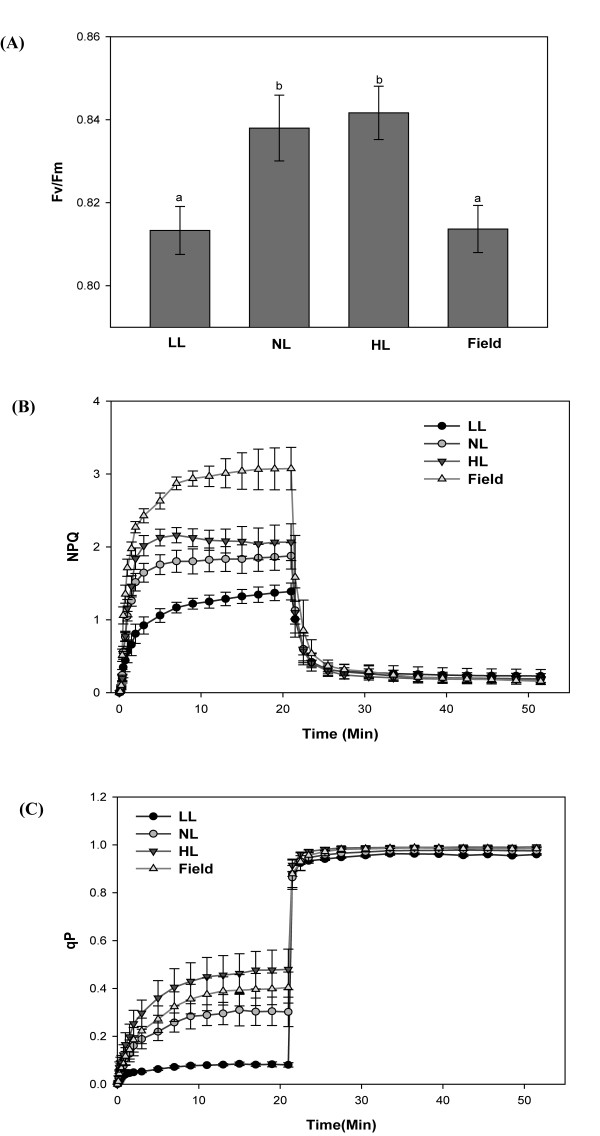
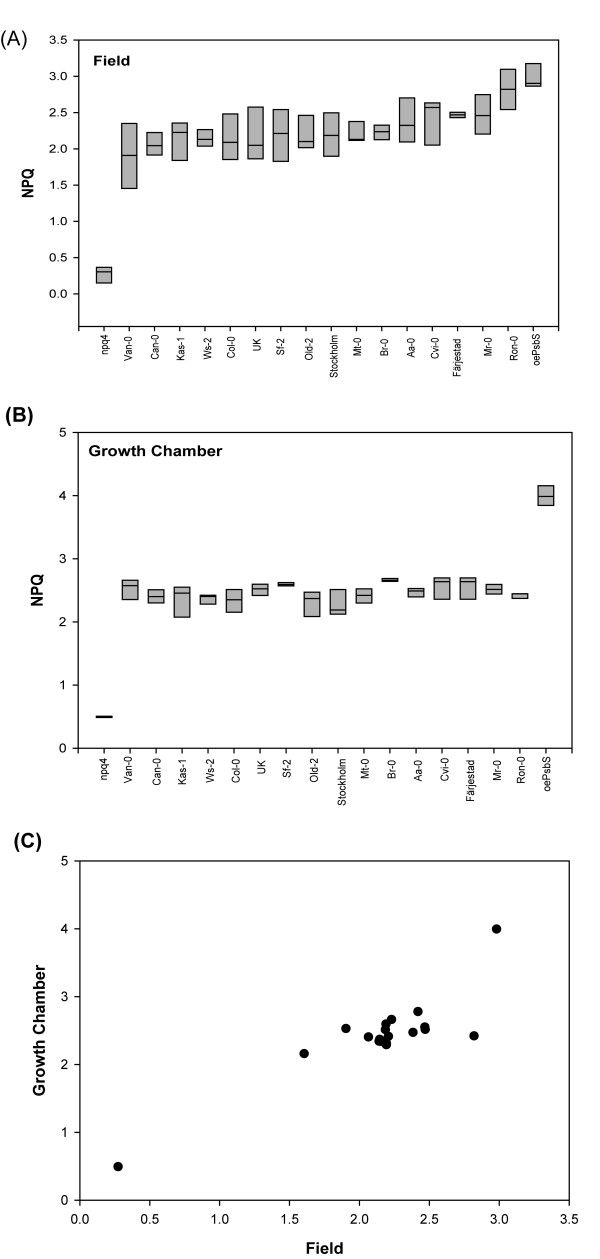
Results: Compared to plants grown under field conditions, the "indoor plants" had larger leaves, modified leaf shapes and longer petioles. Their pigment composition also significantly differed; indoor plants had reduced levels of xanthophyll pigments. In addition, Lhcb1 and Lhcb2 levels were up to three times higher in the indoor plants, but differences in the PSI antenna were much smaller, with only the low-abundance Lhca5 protein showing altered levels. Both isoforms of early-light-induced protein (ELIP) were absent in the indoor plants, and they had less non-photochemical quenching (NPQ). The field-grown plants had a high capacity to perform state transitions. Plants lacking ELIPs did not have reduced growth or seed set rates, but their mortality rates were sometimes higher. NPQ levels between natural accessions grown under different conditions were not correlated.
Conclusion: Our results indicate that comparative analysis of field-grown plants with those grown under artificial conditions is important for a full understanding of plant plasticity and adaptation.
© 2011 Mishra et al; licensee BioMed Central Ltd.
Figures



References
-
- Atkin OK, Loveys BR, Atkinson LJ, Pons TL. Phenotypic plasticity and growth temperature: understanding site-specific variability. J Exp Bot. 2006;57:267–281. - PubMed
-
- Poorter H, Pepin S, Rijkers T, de Jong Y, Evans JR, Körner C. Construction costs, chemical composition and payback time of high- and low-irradiance leaves. J Exp Bot. 2006;57:355–371. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
