

Dissection of the NUP107 nuclear pore subcomplex reveals a novel interaction with spindle assembly checkpoint protein MAD1 in Caenorhabditis elegans
- PMID: 22238360
- PMCID: PMC3290650
- DOI: 10.1091/mbc.E11-11-0927
Dissection of the NUP107 nuclear pore subcomplex reveals a novel interaction with spindle assembly checkpoint protein MAD1 in Caenorhabditis elegans
Abstract
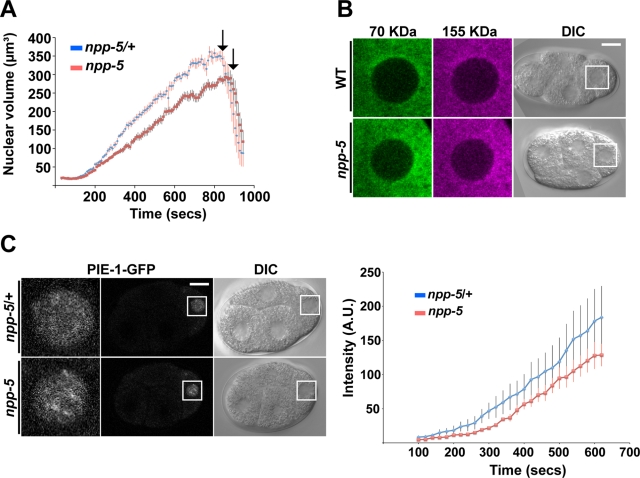
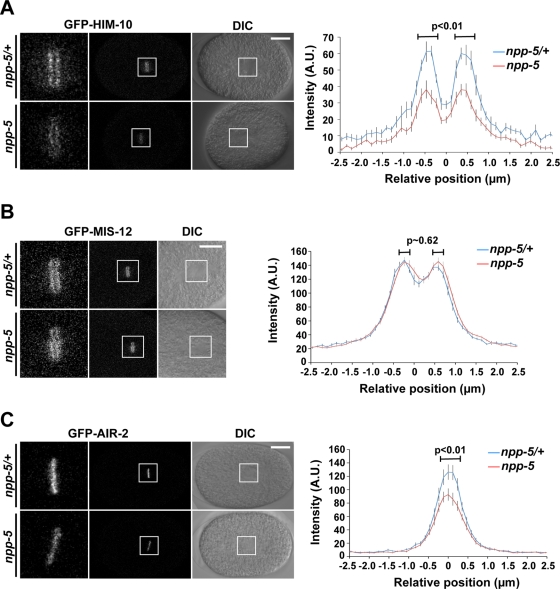
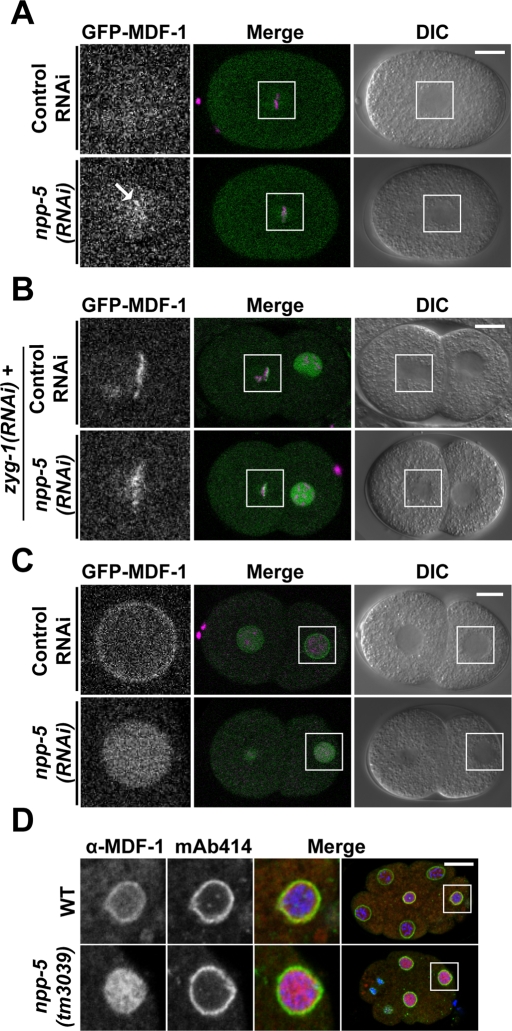
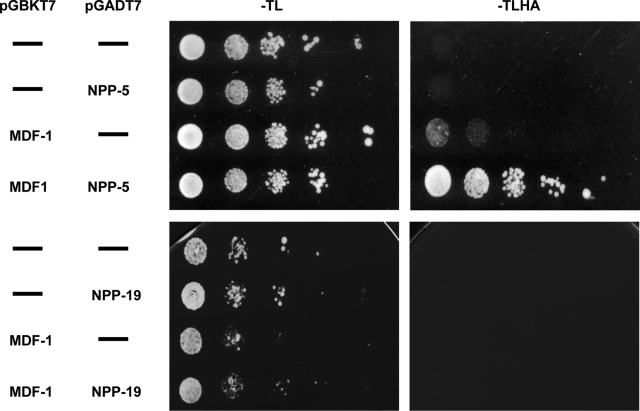
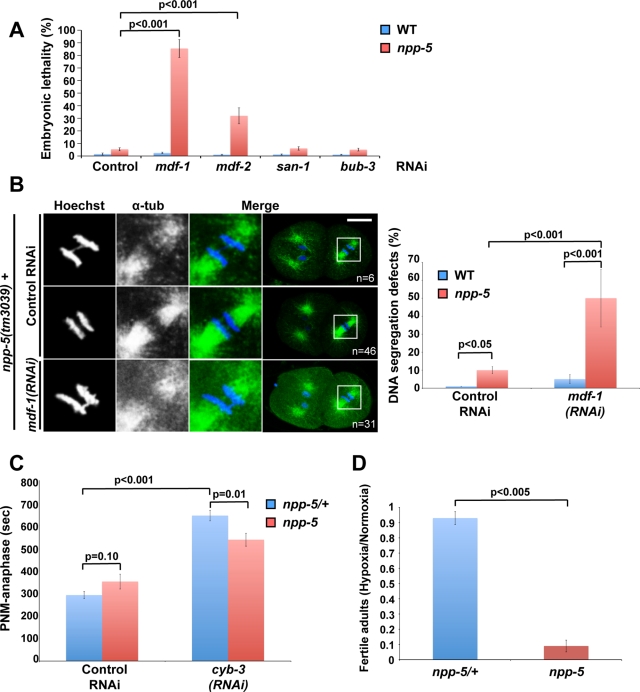
Nuclear pore complexes consist of several subcomplexes. The NUP107 complex is important for nucleocytoplasmic transport, nuclear envelope assembly, and kinetochore function. However, the underlying molecular mechanisms and the roles of individual complex members remain elusive. We report the first description of a genetic disruption of NUP107 in a metazoan. Caenorhabditis elegans NUP107/npp-5 mutants display temperature-dependent lethality. Surprisingly, NPP-5 is dispensable for incorporation of most nucleoporins into nuclear pores and for nuclear protein import. In contrast, NPP-5 is essential for proper kinetochore localization of NUP133/NPP-15, another NUP107 complex member, whereas recruitment of NUP96/NPP-10C and ELYS/MEL-28 is NPP-5 independent. We found that kinetochore protein NUF2/HIM-10 and Aurora B/AIR-2 kinase are less abundant on mitotic chromatin upon NPP-5 depletion. npp-5 mutants are hypersensitive to anoxia, suggesting that the spindle assembly checkpoint (SAC) is compromised. Indeed, NPP-5 interacts genetically and physically with SAC protein MAD1/MDF-1, whose nuclear envelope accumulation requires NPP-5. Thus our results strengthen the emerging connection between nuclear pore proteins and chromosome segregation.
Figures



References
-
- Alber F, et al. The molecular architecture of the nuclear pore complex. Nature. 2007;450:695–701. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
