

Derlin-1 and UBXD8 are engaged in dislocation and degradation of lipidated ApoB-100 at lipid droplets
- PMID: 22238364
- PMCID: PMC3290640
- DOI: 10.1091/mbc.E11-11-0950
Derlin-1 and UBXD8 are engaged in dislocation and degradation of lipidated ApoB-100 at lipid droplets
Abstract
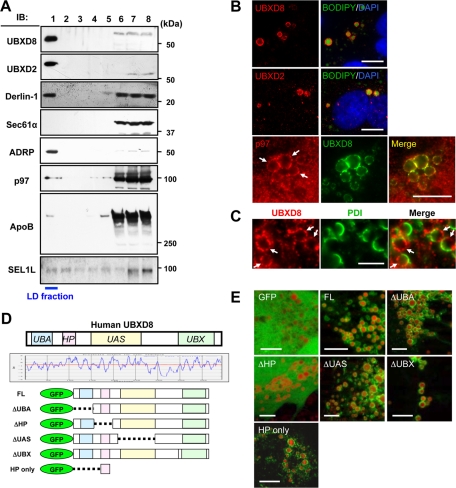
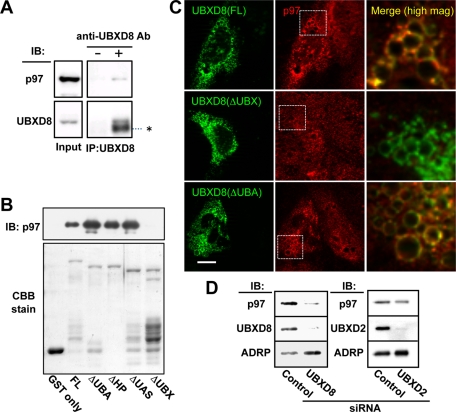
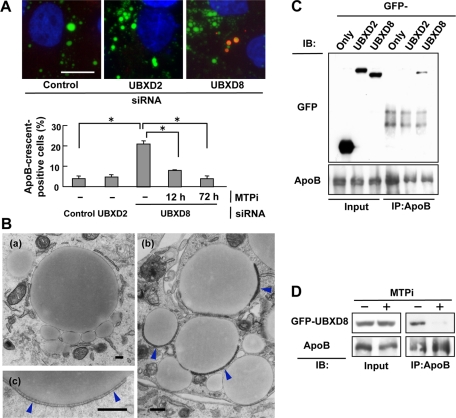
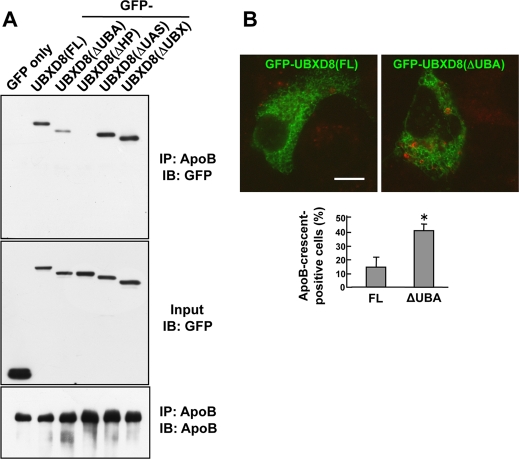
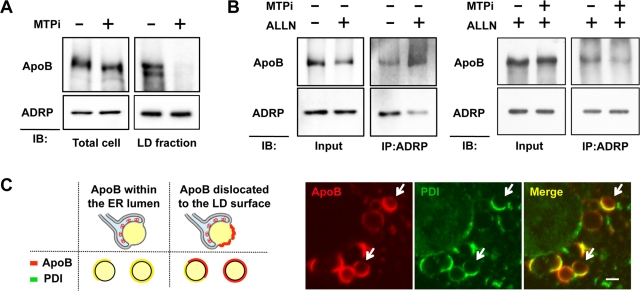
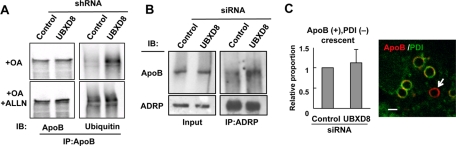
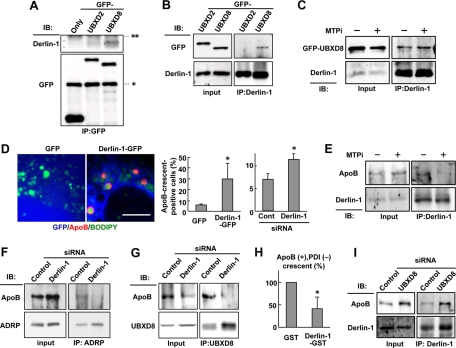
Apolipoprotein B-100 (ApoB) is the principal component of very low density lipoprotein. Poorly lipidated nascent ApoB is extracted from the Sec61 translocon and degraded by proteasomes. ApoB lipidated in the endoplasmic reticulum (ER) lumen is also subjected to proteasomal degradation, but where and how it dislocates to the cytoplasm remain unknown. In the present study, we demonstrate that ApoB after lipidation is dislocated to the cytoplasmic surface of lipid droplets (LDs) and accumulates as ubiquitinated ApoB in Huh7 cells. Depletion of UBXD8, which is almost confined to LDs in this cell type, decreases recruitment of p97 to LDs and causes an increase of both ubiquitinated ApoB on the LD surface and lipidated ApoB in the ER lumen. In contrast, abrogation of Derlin-1 function induces an accumulation of lipidated ApoB in the ER lumen but does not increase ubiquitinated ApoB on the LD surface. UBXD8 and Derlin-1 bind with each other and with lipidated ApoB and show colocalization around LDs. These results indicate that ApoB after lipidation is dislocated from the ER lumen to the LD surface for proteasomal degradation and that Derlin-1 and UBXD8 are engaged in the predislocation and postdislocation steps, respectively.
Figures

References
-
- Bagola K, Mehnert M, Jarosch E, Sommer T. Protein dislocation from the ER. Biochim Biophys Acta. 2011;1808:925–936. - PubMed
-
- Bartz R, Zehmer JK, Zhu M, Chen Y, Serrero G, Zhao Y, Liu P. Dynamic activity of lipid droplets: protein phosphorylation and GTP-mediated protein translocation. J Proteome Res. 2007;6:3256–3265. - PubMed
-
- Brasaemle DL, Barber T, Kimmel AR, Londos C. Post-translational regulation of perilipin expression. Stabilization by stored intracellular neutral lipids. J Biol Chem. 1997;272:9378–9387. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
