

Classification of protein functional surfaces using structural characteristics
- PMID: 22238424
- PMCID: PMC3268291
- DOI: 10.1073/pnas.1119684109
Classification of protein functional surfaces using structural characteristics
Abstract
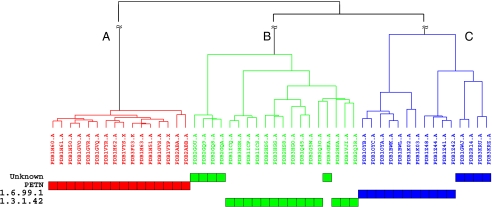
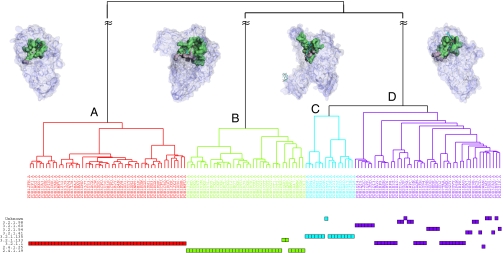
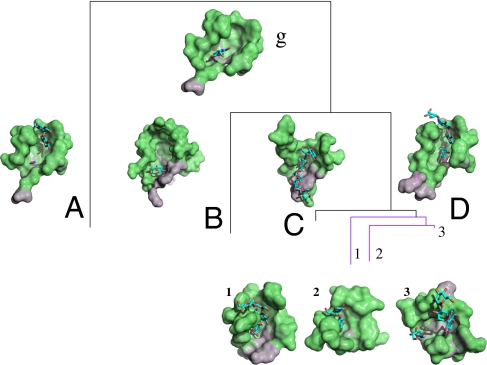
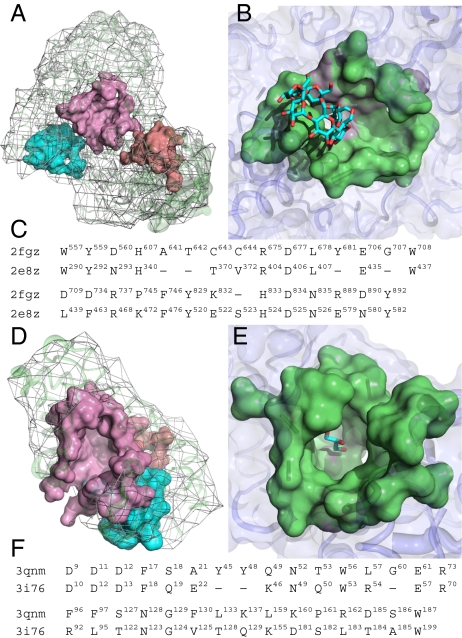
Protein structure and function are closely related, especially in functional surfaces, which are local spatial regions that perform the biological functions. Also, protein structures tend to evolve more slowly than amino acid sequences. We have therefore developed a method to classify proteins using the structures of functional surfaces; we call it protein surface classification (PSC). PSC may reflect functional relationships among proteins and may detect evolutionary relationships among highly divergent sequences. We focused on the surfaces of ligand-bound regions because they represent well-defined structures. Specifically, we used structural attributes to measure similarities between binding surfaces and constructed a PSC library of ~2,000 binding surface types from the bound forms. Using flavin mononucleotide-binding proteins and glycosidases as examples, we show how the evolutionary position of an uncharacterized protein can be defined and its function inferred from the characterized members of the same surface subtype. We found that proteins with the same enzyme nomenclature may be divided into subtypes and that two proteins in the same CATH (Class, Architecture, Topology, Homologous superfamily) fold may belong to two different surface types. In conclusion, our approach complements the sequence-based and fold-domain classifications and has the advantage of associating the shape of a protein with its biological function. As an expandable library, PSC provides a resource of spatial patterns for studying the evolution of protein structure and function.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
PSC: protein surface classification.Nucleic Acids Res. 2012 Jul;40(Web Server issue):W435-9. doi: 10.1093/nar/gks495. Epub 2012 Jun 4. Nucleic Acids Res. 2012. PMID: 22669905 Free PMC article.
-
CATH--a hierarchic classification of protein domain structures.Structure. 1997 Aug 15;5(8):1093-108. doi: 10.1016/s0969-2126(97)00260-8. Structure. 1997. PMID: 9309224
-
Inferring functional relationships of proteins from local sequence and spatial surface patterns.J Mol Biol. 2003 Sep 12;332(2):505-26. doi: 10.1016/s0022-2836(03)00882-9. J Mol Biol. 2003. PMID: 12948498
-
Protein surface similarities: a survey of methods to describe and compare protein surfaces.Cell Mol Life Sci. 2000 Dec;57(13-14):1970-7. doi: 10.1007/PL00000677. Cell Mol Life Sci. 2000. PMID: 11215522 Free PMC article. Review.
-
The family feud: do proteins with similar structures fold via the same pathway?Curr Opin Struct Biol. 2005 Feb;15(1):42-9. doi: 10.1016/j.sbi.2005.01.011. Curr Opin Struct Biol. 2005. PMID: 15718132 Review.
Cited by
-
PSC: protein surface classification.Nucleic Acids Res. 2012 Jul;40(Web Server issue):W435-9. doi: 10.1093/nar/gks495. Epub 2012 Jun 4. Nucleic Acids Res. 2012. PMID: 22669905 Free PMC article.
-
Measuring the shapes of macromolecules - and why it matters.Comput Struct Biotechnol J. 2013 Dec 9;8:e201309001. doi: 10.5936/csbj.201309001. eCollection 2013. Comput Struct Biotechnol J. 2013. PMID: 24688748 Free PMC article. Review.
-
Functional consequences of somatic mutations in cancer using protein pocket-based prioritization approach.Genome Med. 2014 Oct 14;6(10):81. doi: 10.1186/s13073-014-0081-7. eCollection 2014. Genome Med. 2014. PMID: 25360158 Free PMC article.
-
Molecular Basis of ABHD5 Lipolysis Activation.Sci Rep. 2017 Feb 17;7:42589. doi: 10.1038/srep42589. Sci Rep. 2017. PMID: 28211464 Free PMC article.
-
The value of protein structure classification information-Surveying the scientific literature.Proteins. 2015 Nov;83(11):2025-38. doi: 10.1002/prot.24915. Epub 2015 Sep 19. Proteins. 2015. PMID: 26313554 Free PMC article.
References
-
- Orengo CA, et al. CATH—A hierarchic classification of protein domain structures. Structure. 1997;5:1093–1108. - PubMed
-
- Murzin AG, Brenner SE, Hubbard T, Chothia C. SCOP: A structural classification of proteins database for the investigation of sequences and structures. J Mol Biol. 1995;247:536–540. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
