

A loss-of-function mutation in the nucleoporin AtNUP160 indicates that normal auxin signalling is required for a proper ethylene response in Arabidopsis
- PMID: 22238449
- PMCID: PMC3295400
- DOI: 10.1093/jxb/err424
A loss-of-function mutation in the nucleoporin AtNUP160 indicates that normal auxin signalling is required for a proper ethylene response in Arabidopsis
Abstract
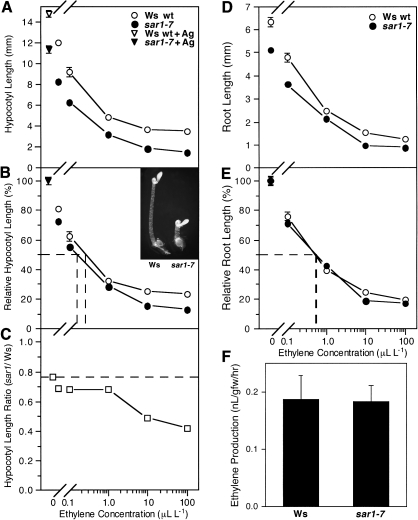
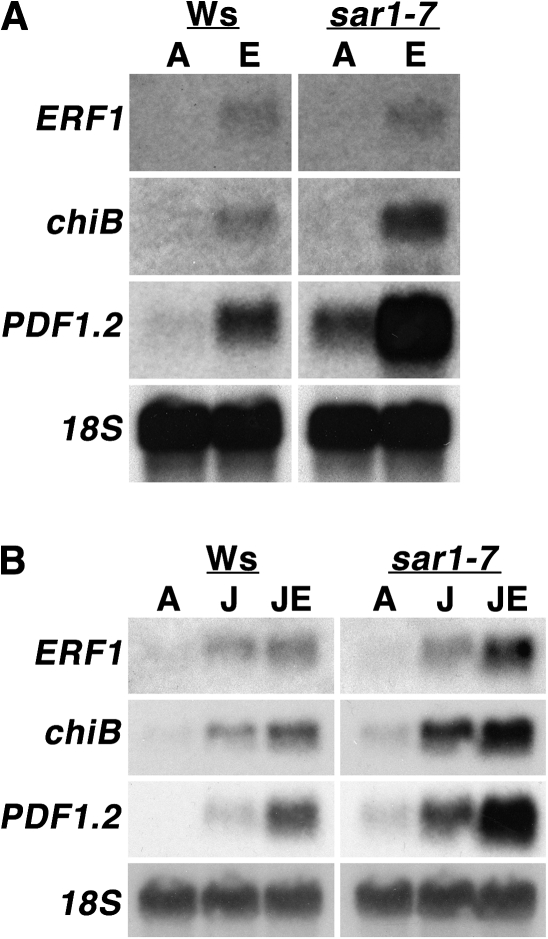
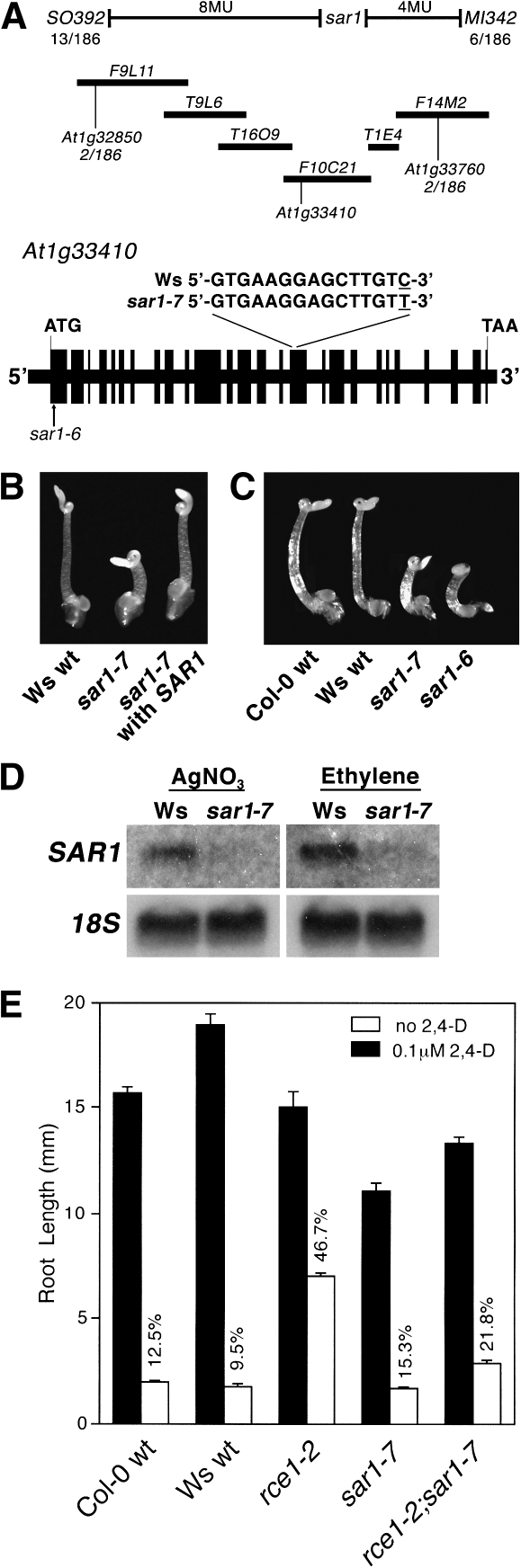
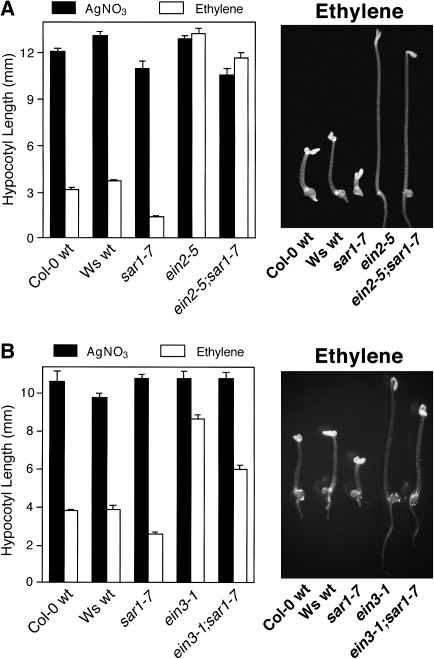
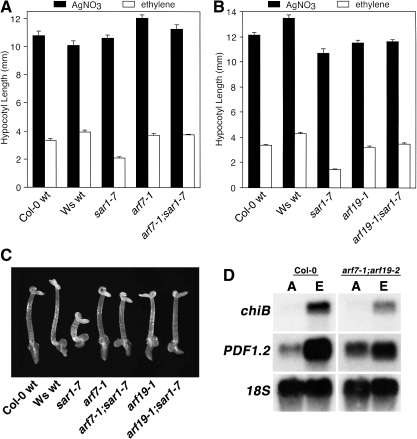
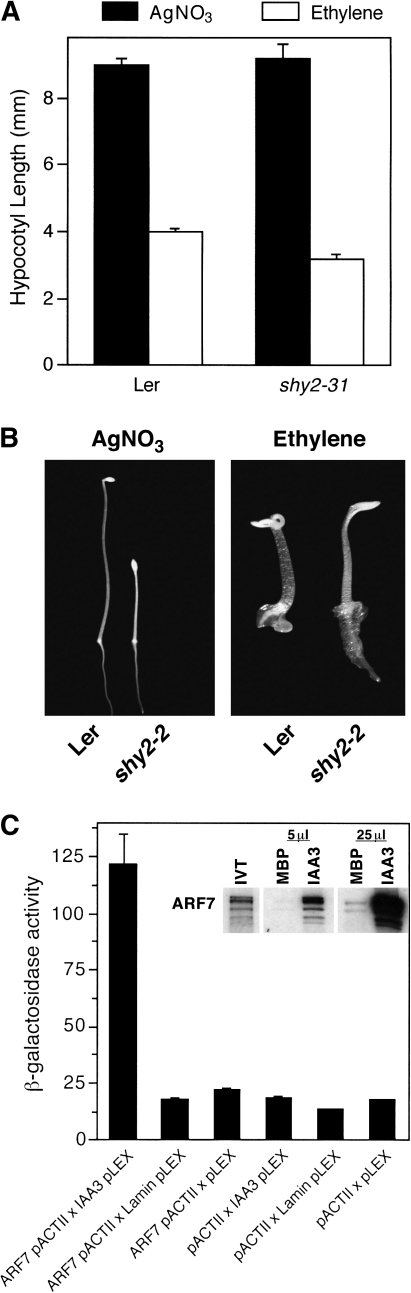
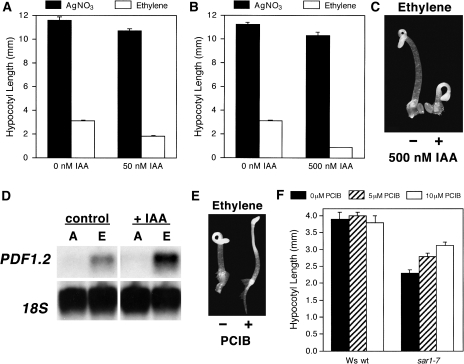
As part of a continuing effort to elucidate mechanisms that regulate the magnitude of ethylene signalling, an Arabidopsis mutant with an enhanced ethylene response was identified. Subsequent characterization of this loss-of-function mutant revealed severe hypocotyl shortening in the presence of saturating ethylene along with increased expression in leaves of a subset of ethylene-responsive genes. It was subsequently determined by map-based cloning that the mutant (sar1-7) represents a loss-of-function mutation in the previously described nucleoporin AtNUP160 (At1g33410, SAR1). In support of previously reported results, the sar1-7 mutant partially restored auxin responsiveness to roots of an rce1 loss-of-function mutant, indicating that AtNUP160/SAR1 is required for proper expression of factors responsible for the repression of auxin signalling. Analysis of arf7-1/sar1-7 and arf19-1/sar1-7 double mutants revealed that mutations affecting either ARF7 or ARF19 function almost fully blocked manifestation of the sar1-7-dependent ethylene hypersensitivity phenotype, suggesting that ARF7- and ARF19-mediated auxin signalling is responsible for regulating the magnitude of and/or competence for the ethylene response in Arabidopsis etiolated hypocotyls. Consistent with this, addition of auxin to ethylene-treated seedlings resulted in severe hypocotyl shortening, reminiscent of that seen for other eer (enhanced ethylene response) mutants, suggesting that auxin functions in part synergistically with ethylene to control hypocotyl elongation and other ethylene-dependent phenomena.
Figures
References
-
- Alonso JM, Hirayama T, Roman G, Nourizadeh S, Ecker JR. EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science. 1999;284:2148–2152. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
