

Convergent differential regulation of parvalbumin in the brains of vocal learners
- PMID: 22238614
- PMCID: PMC3253077
- DOI: 10.1371/journal.pone.0029457
Convergent differential regulation of parvalbumin in the brains of vocal learners
Abstract
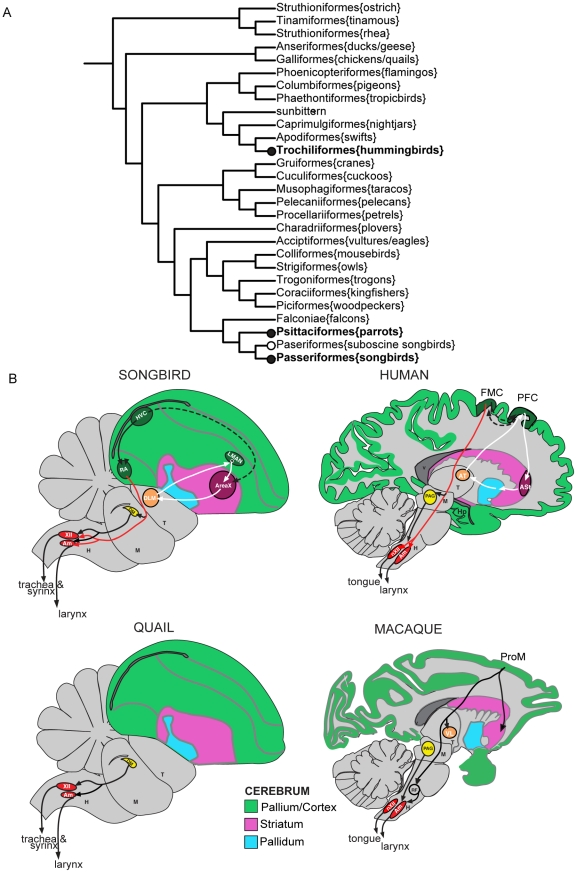
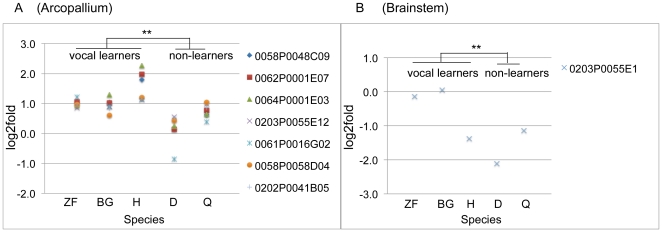
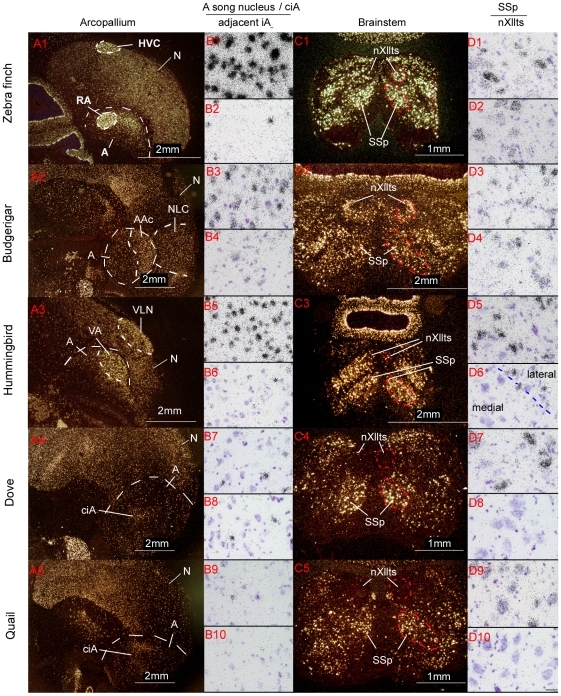
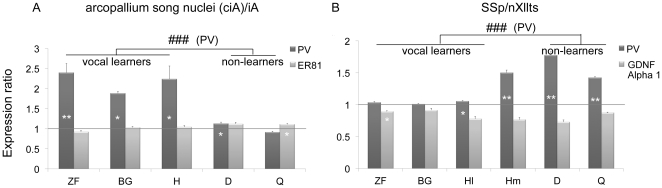
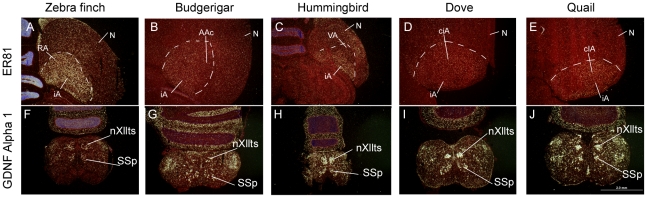
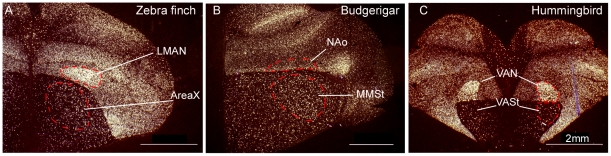
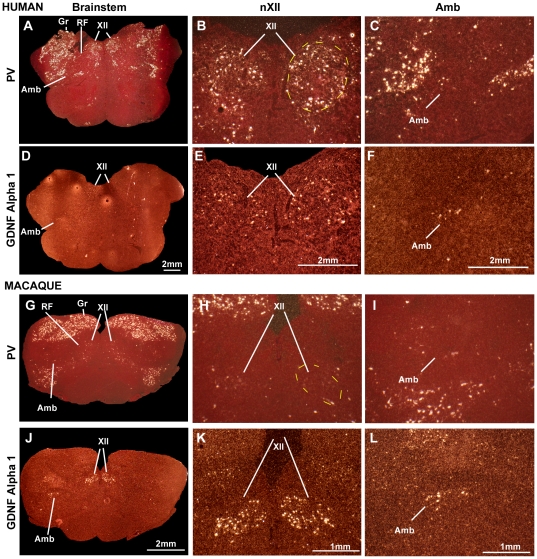
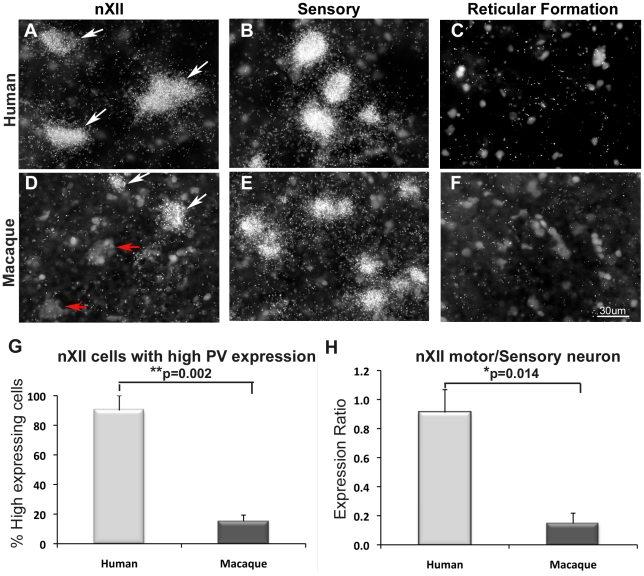
Spoken language and learned song are complex communication behaviors found in only a few species, including humans and three groups of distantly related birds--songbirds, parrots, and hummingbirds. Despite their large phylogenetic distances, these vocal learners show convergent behaviors and associated brain pathways for vocal communication. However, it is not clear whether this behavioral and anatomical convergence is associated with molecular convergence. Here we used oligo microarrays to screen for genes differentially regulated in brain nuclei necessary for producing learned vocalizations relative to adjacent brain areas that control other behaviors in avian vocal learners versus vocal non-learners. A top candidate gene in our screen was a calcium-binding protein, parvalbumin (PV). In situ hybridization verification revealed that PV was expressed significantly higher throughout the song motor pathway, including brainstem vocal motor neurons relative to the surrounding brain regions of all distantly related avian vocal learners. This differential expression was specific to PV and vocal learners, as it was not found in avian vocal non-learners nor for control genes in learners and non-learners. Similar to the vocal learning birds, higher PV up-regulation was found in the brainstem tongue motor neurons used for speech production in humans relative to a non-human primate, macaques. These results suggest repeated convergent evolution of differential PV up-regulation in the brains of vocal learners separated by more than 65-300 million years from a common ancestor and that the specialized behaviors of learned song and speech may require extra calcium buffering and signaling.
Conflict of interest statement
Figures
Similar articles
-
Differential expression of glutamate receptors in avian neural pathways for learned vocalization.J Comp Neurol. 2004 Aug 9;476(1):44-64. doi: 10.1002/cne.20201. J Comp Neurol. 2004. PMID: 15236466 Free PMC article.
-
Comparative analysis of gene expressions among avian brains: a molecular approach to the evolution of vocal learning.Brain Res Bull. 2008 Mar 18;75(2-4):474-9. doi: 10.1016/j.brainresbull.2007.10.045. Epub 2007 Nov 20. Brain Res Bull. 2008. PMID: 18331917
-
Evolution and diversity in avian vocal system: an Evo-Devo model from the morphological and behavioral perspectives.Dev Growth Differ. 2009 Apr;51(3):355-67. doi: 10.1111/j.1440-169X.2009.01091.x. Epub 2008 Feb 16. Dev Growth Differ. 2009. PMID: 19222524 Review.
-
Genetic components of vocal learning.Ann N Y Acad Sci. 2004 Jun;1016:325-47. doi: 10.1196/annals.1298.032. Ann N Y Acad Sci. 2004. PMID: 15313783 Review.
-
FoxP2 expression in avian vocal learners and non-learners.J Neurosci. 2004 Mar 31;24(13):3164-75. doi: 10.1523/JNEUROSCI.4369-03.2004. J Neurosci. 2004. PMID: 15056696 Free PMC article.
Cited by
-
Specialized motor-driven dusp1 expression in the song systems of multiple lineages of vocal learning birds.PLoS One. 2012;7(8):e42173. doi: 10.1371/journal.pone.0042173. Epub 2012 Aug 2. PLoS One. 2012. PMID: 22876306 Free PMC article.
-
Dynamic gene expression in the song system of zebra finches during the song learning period.Dev Neurobiol. 2015 Dec;75(12):1315-38. doi: 10.1002/dneu.22286. Epub 2015 Mar 20. Dev Neurobiol. 2015. PMID: 25787707 Free PMC article.
-
The shape of the human language-ready brain.Front Psychol. 2014 Apr 4;5:282. doi: 10.3389/fpsyg.2014.00282. eCollection 2014. Front Psychol. 2014. PMID: 24772099 Free PMC article.
-
Cross-species conservation in the regulation of parvalbumin by perineuronal nets.Front Neural Circuits. 2023 Dec 19;17:1297643. doi: 10.3389/fncir.2023.1297643. eCollection 2023. Front Neural Circuits. 2023. PMID: 38179221 Free PMC article.
-
Neuroestrogens rapidly shape auditory circuits to support communication learning and perception: Evidence from songbirds.Horm Behav. 2018 Aug;104:77-87. doi: 10.1016/j.yhbeh.2018.03.007. Epub 2018 Mar 30. Horm Behav. 2018. PMID: 29555375 Free PMC article. Review.
References
-
- Janik VM, Salater PB. Vocal learning in mammals. Adv Study Behav. 1997;26:59–99.
-
- Doupe AJ, Kuhl PK. Birdsong and human speech: common themes and mechanisms. Annu Rev Neurosci. 1999;22:567–631. - PubMed
-
- Hauser MD, Chomsky N, Fitch WT. The faculty of language: what is it, who has it, and how did it evolve? Science. 2002;298:1569–1579. - PubMed
-
- Nottebohm F. The origins of vocal learning. The American Naturalist. 1972;106:116–140.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
