

doi: 10.1021/ja210496n.
Epub 2012 Jan 12.
A molecular mechanism for modulating plasma Zn speciation by fatty acids
Affiliations
- PMID: 22239162
- PMCID: PMC3285120
- DOI: 10.1021/ja210496n
Item in Clipboard
A molecular mechanism for modulating plasma Zn speciation by fatty acids
J Am Chem Soc.
.
Abstract
Albumin transports both fatty acids and zinc in plasma. Competitive binding studied by isothermal titration calorimetry revealed that physiologically relevant levels of fatty acids modulate the Zn-binding capacity of albumin, with far-reaching implications for biological zinc speciation. The molecular mechanism for this effect is likely due to a large conformational change elicited by fatty acid binding to a high-affinity interdomain site that disrupts at least one Zn site. Albumin may be a molecular device to "translate" certain aspects of the organismal energy state into global zinc signals.
© 2012 American Chemical Society
Figures
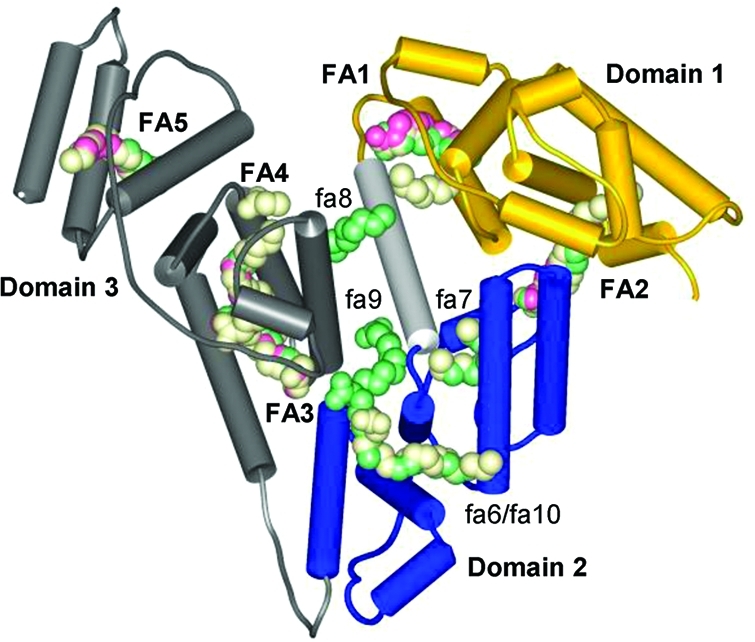
Domain structure of albumin
and fatty acid binding sites. Overlaid
structures with PDB codes: 1bj5, HSA with five myristates, pink (the protein backbone
is also shown); 1e7e, HSA with 10 decanoates, green; 1gnj, HSA with seven arachidonates, light-yellow. FA1–5, major sites; fa6–10, minor
sites.
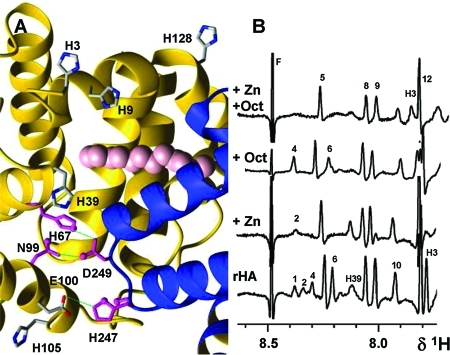
Binding events at the domain I/II interface
affect several His
Hε1 resonances. (A) Interface with the disrupted Zn-binding
site A (magenta) and interdomain His residues likely to be affected
by fatty acid binding in site FA2. The 11 resolved carbons of Myr2
in PDB entry 1bj5 are shown as pink spheres. (B) Effects of 1 molar equiv of Zn and
5 molar equiv of OCT on His Hε1 resonances. Peaks 1 and 4 are
assigned to His67 and His247; full titration
data are shown in Figure S3.
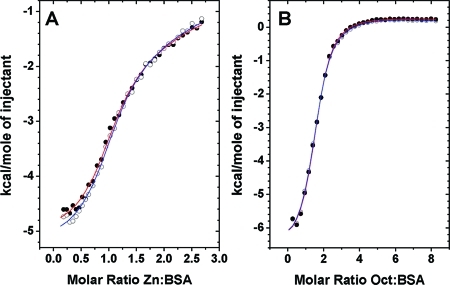
Competitive binding of
Zn and OCT to BSA studied by ITC. (A) Zn
binding to BSA (25 μM) in the presence and absence of 5 molar
equiv of OCT. (B) OCT binding to BSA and Zn1BSA. Open circles
with blue fits correspond to binary systems and filled circles with
red fits to ternary systems. In each case, 34 injections of 8 μL
of 333 μM ligand (Zn or OCT) were delivered over 16 s with 240
s between injections for complete equilibration.
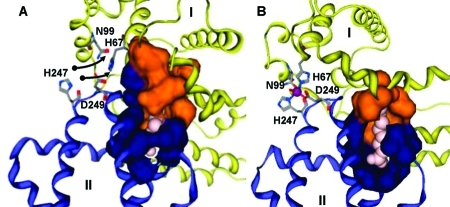
Different binding modes for (A) medium- and (B) short-chain
fatty
acids in site FA2 on HSA. Fatty acid molecules are shown in pink.
The colored surfaces represent Analytical Connolly surfaces of the
residues forming the binding pocket. In both models, the carboxylate
headgroup interacts with R257, and the hydrophobic half-pocket in
domain II (blue) is formed by residues L250, L251, A254, A258, L283,
and L284. (A) HSA with bound MYR, based on PDB entry 1bj5. Three C atoms have
been added to the C11 chain resolved in the X-ray structure. Domain
I (orange and yellow) contributes to the fatty acid binding site an
extended half-pocket comprising residues R10, L14, F19, L22, V23,
A26, L66, and Y150. The complete pocket can be formed only if the
zinc site (labeled residues) is disrupted. (B) HSA with OCT and Zn2+ (purple) bound simultaneously. OCT is short enough to be
accommodated predominantly in the domain II pocket. Hydrophobic residues
L14, F19, L22, and L155 form a new half-pocket without disrupting
the zinc site.
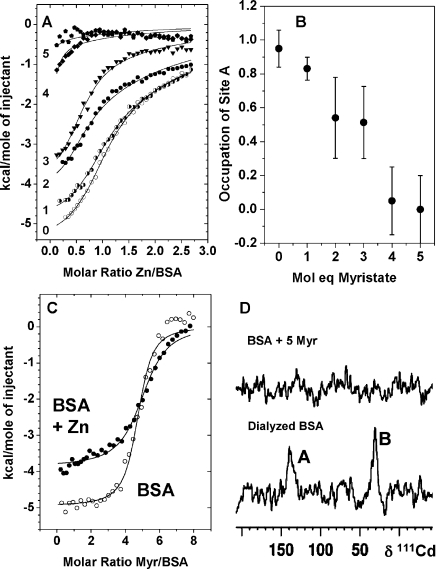
Competitive binding of
metals and MYR to BSA. (A) Effect of increasing
amounts of MYR on the zinc-binding capacity of BSA. ITC curves for
titrations of 333 μM Zn2+ into 25 μM BSA in
the presence and absence of varying amounts (0–5 molar equiv)
of MYR in 50 mM Tris/50 mM NaCl (pH 7.2). The fits (Figure S8) allowed estimates of the ratio of site A availability,
as shown in (B). A clear downward trend was observed. A 4:1 MYR:Zn
molar ratio suppressed occupation of site A almost completely. A second
binding site was also affected by fatty acid binding (see D). (C)
ITC curves for titrations of 500 μM MYR into 12.5 μM BSA
or Zn1BSA. These titrations were carried out in H2O because of the insufficient solubility of MYR in Tris buffer. The
fits (Figure S9) correspond to a model
with one set of binding sites to estimate the stoichiometry for the
highest-affinity sites. This equaled 5.0 ± 0.3, and the average
log K was 6.3 ± 0.4 in the absence and 5.9 ±
0.4 in the presence of Zn2+. More complex fits were possible,
but the data were insufficient to justify them. (D) 111Cd NMR spectra of Cd2BSA recorded in the absence and presence
of 5 molar equiv of MYR. Peaks A and B were both suppressed by MYR.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
