

Autophagy releases lipid that promotes fibrogenesis by activated hepatic stellate cells in mice and in human tissues
- PMID: 22240484
- PMCID: PMC3439519
- DOI: 10.1053/j.gastro.2011.12.044
Autophagy releases lipid that promotes fibrogenesis by activated hepatic stellate cells in mice and in human tissues
Abstract
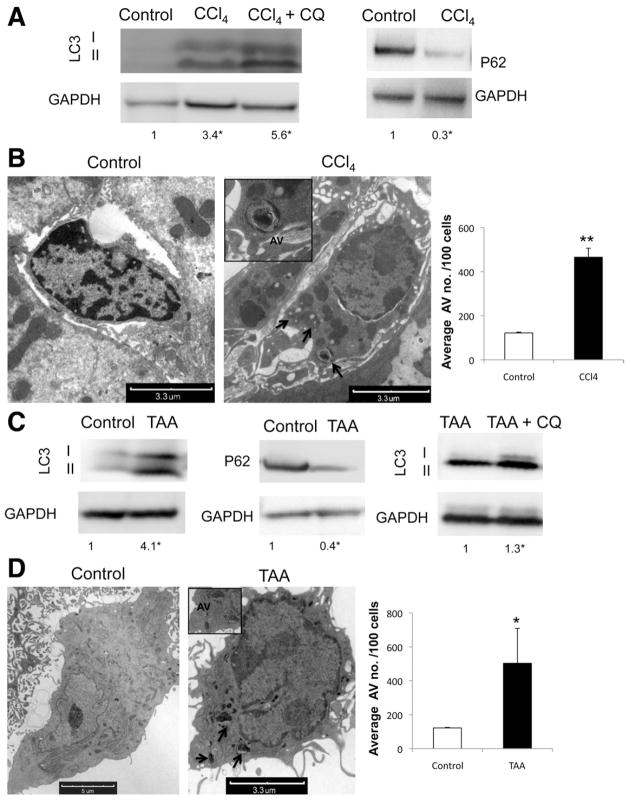
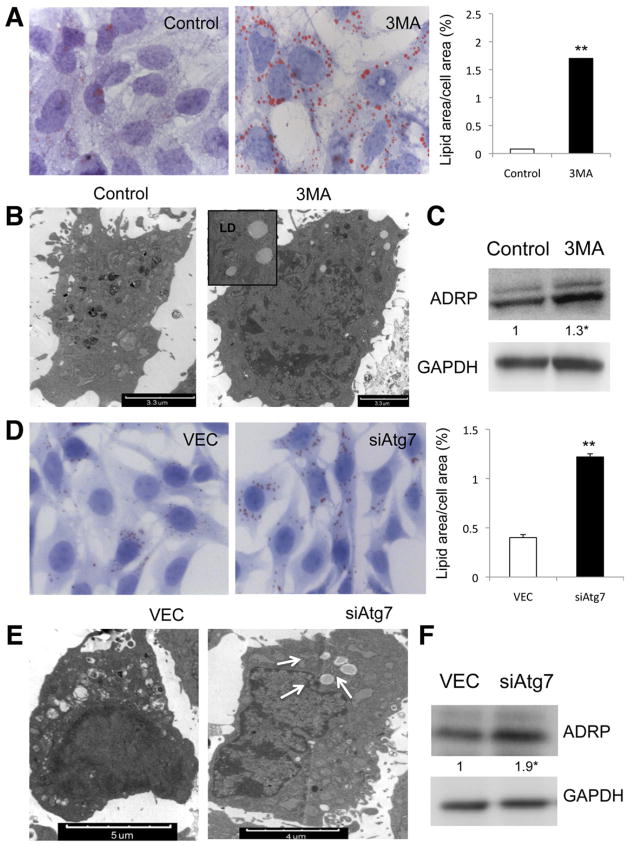
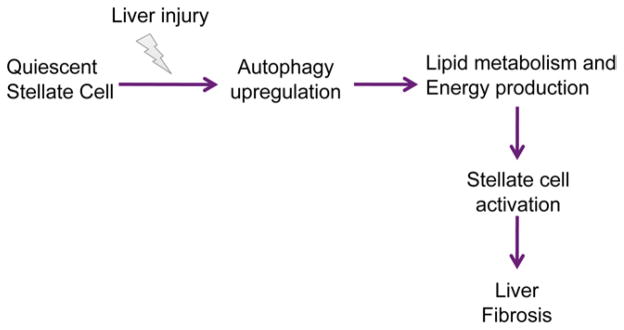
Background & aims: The pathogenesis of liver fibrosis involves activation of hepatic stellate cells, which is associated with depletion of intracellular lipid droplets. When hepatocytes undergo autophagy, intracellular lipids are degraded in lysosomes. We investigated whether autophagy also promotes loss of lipids in hepatic stellate cells to provide energy for their activation and extended these findings to other fibrogenic cells.
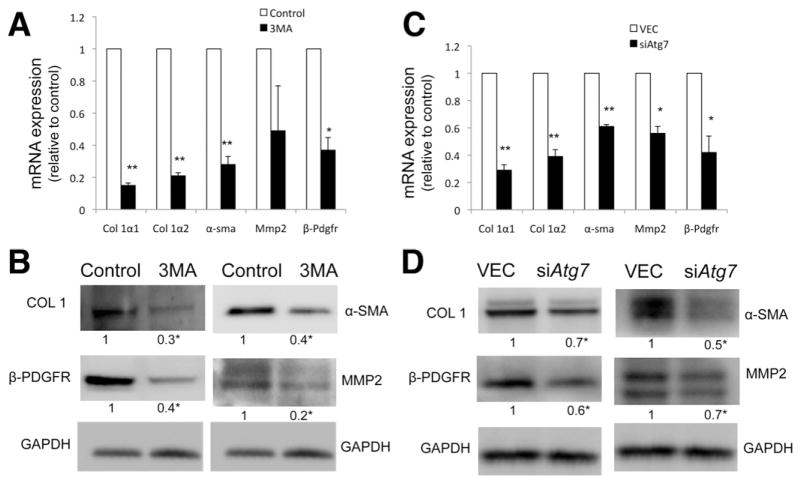
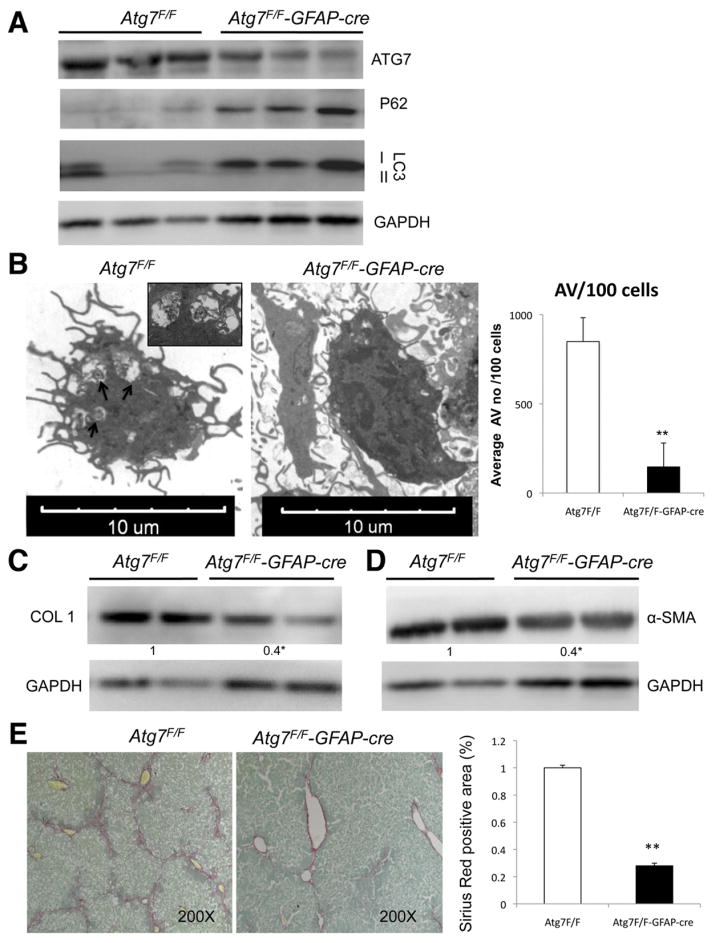
Methods: We analyzed hepatic stellate cells from C57BL/6 wild-type, Atg7(F/F), and Atg7(F/F)-GFAP-Cre mice, as well as the mouse stellate cell line JS1. Fibrosis was induced in mice using CCl(4) or thioacetamide (TAA); liver tissues and stellate cells were analyzed. Autophagy was blocked in fibrogenic cells from liver and other tissues using small interfering RNAs against Atg5 or Atg7 and chemical antagonists. Human pulmonary fibroblasts were isolated from samples of lung tissue from patients with idiopathic pulmonary fibrosis or from healthy donors.
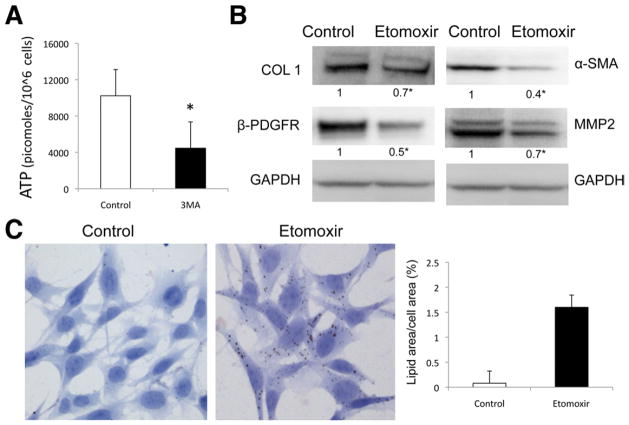
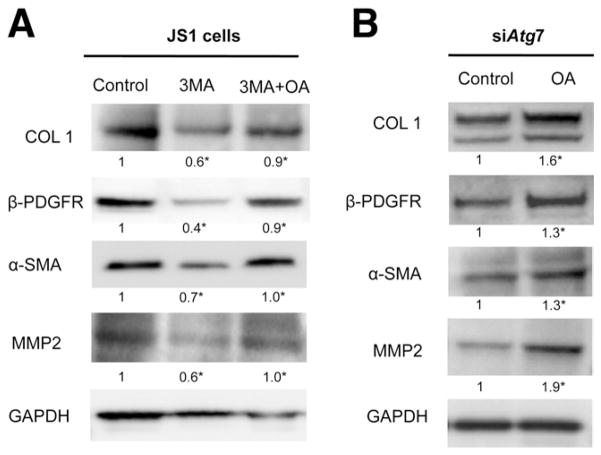
Results: In mice, induction of liver injury with CCl(4) or TAA increased levels of autophagy. We also observed features of autophagy in activated stellate cells within injured human liver tissue. Loss of autophagic function in cultured mouse stellate cells and in mice following injury reduced fibrogenesis and matrix accumulation; this effect was partially overcome by providing oleic acid as an energy substrate. Autophagy also regulated expression of fibrogenic genes in embryonic, lung, and renal fibroblasts.
Conclusions: Autophagy of activated stellate cells is required for hepatic fibrogenesis in mice. Selective reduction of autophagic activity in fibrogenic cells in liver and other tissues might be used to treat patients with fibrotic diseases.
Copyright © 2012 AGA Institute. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interest
The authors disclose no conflicts.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK056621/DK/NIDDK NIH HHS/United States
- R01 DK044234/DK/NIDDK NIH HHS/United States
- KO5AA018408/AA/NIAAA NIH HHS/United States
- R01DK044234/DK/NIDDK NIH HHS/United States
- R01NS060123/NS/NINDS NIH HHS/United States
- 5T32GM007280/GM/NIGMS NIH HHS/United States
- P20AA017067/AA/NIAAA NIH HHS/United States
- K05 AA018408/AA/NIAAA NIH HHS/United States
- P20 AA017067/AA/NIAAA NIH HHS/United States
- R01 AA020709/AA/NIAAA NIH HHS/United States
- T32 GM007280/GM/NIGMS NIH HHS/United States
- R01DK061498/DK/NIDDK NIH HHS/United States
- R01 DK037340/DK/NIDDK NIH HHS/United States
- R01 NS060123/NS/NINDS NIH HHS/United States
- R01 DK061498/DK/NIDDK NIH HHS/United States
- R01DK56621/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
