

Structural bias in T4 RNA ligase-mediated 3'-adapter ligation
- PMID: 22241775
- PMCID: PMC3326334
- DOI: 10.1093/nar/gkr1263
Structural bias in T4 RNA ligase-mediated 3'-adapter ligation
Abstract
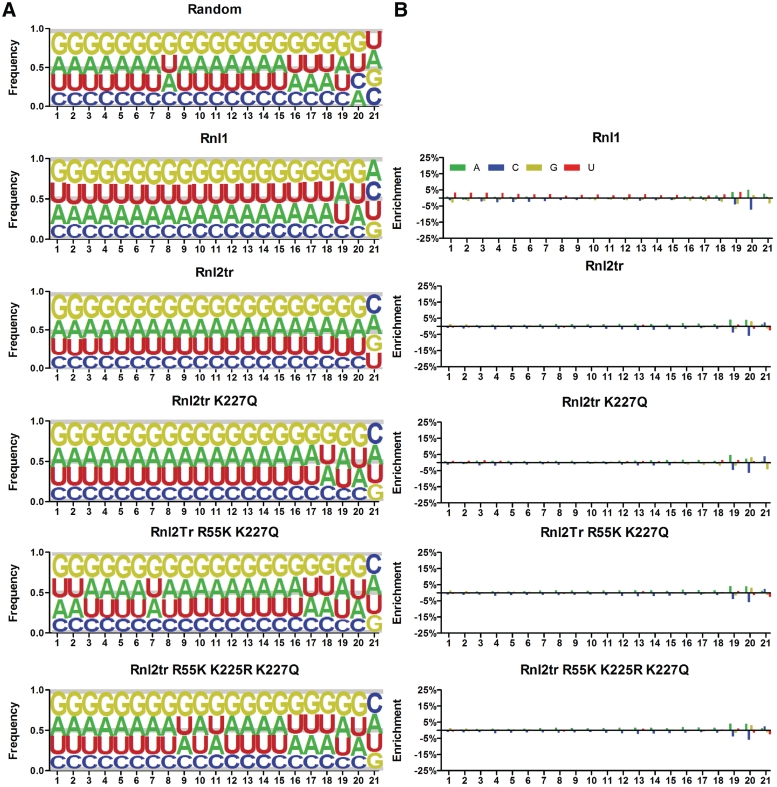
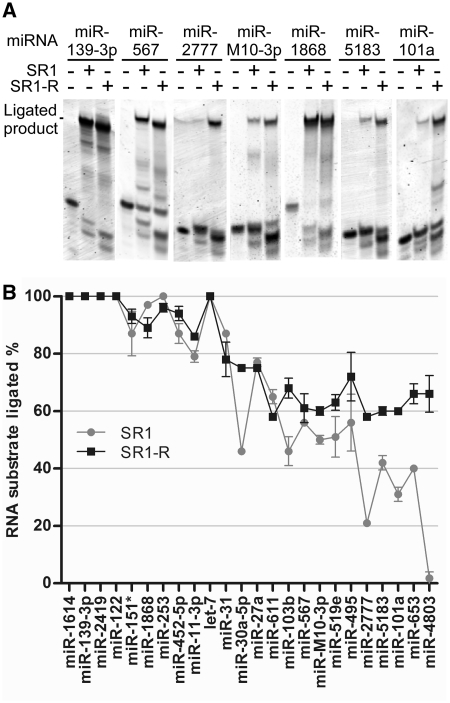
T4 RNA ligases are commonly used to attach adapters to RNAs, but large differences in ligation efficiency make detection and quantitation problematic. We developed a ligation selection strategy using random RNAs in combination with high-throughput sequencing to gain insight into the differences in efficiency of ligating pre-adenylated DNA adapters to RNA 3'-ends. After analyzing biases in RNA sequence, secondary structure and RNA-adapter cofold structure, we conclude that T4 RNA ligases do not show significant primary sequence preference in RNA substrates, but are biased against structural features within RNAs and adapters. Specifically, RNAs with less than three unstructured nucleotides at the 3'-end and RNAs that are predicted to cofold with an adapter in unfavorable structures are likely to be poorly ligated. The effect of RNA-adapter cofold structures on ligation is supported by experiments where the ligation efficiency of specific miRNAs was changed by designing adapters to alter cofold structure. In addition, we show that using adapters with randomized regions results in higher ligation efficiency and reduced ligation bias. We propose that using randomized adapters may improve RNA representation in experiments that include a 3'-adapter ligation step.
Figures







References
-
- Cranston JW, Silber R, Malathi VG, Hurwitz J. Studies on ribonucleic acid ligase. Characterization of an adenosine triphosphate-inorganic pyrophosphate exchange reaction and demonstration of an enzyme-adenylate complex with T4 bacteriophage-induced enzyme. J. Biol. Chem. 1974;249:7447–7456. - PubMed
-
- Sugino A, Snoper TJ, Cozzarelli NR. Bacteriophage T4 RNA ligase. Reaction intermediates and interaction of substrates. J. Biol. Chem. 1977;252:1732–1738. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
