

Eight RGS and RGS-like proteins orchestrate growth, differentiation, and pathogenicity of Magnaporthe oryzae
- PMID: 22241981
- PMCID: PMC3248559
- DOI: 10.1371/journal.ppat.1002450
Eight RGS and RGS-like proteins orchestrate growth, differentiation, and pathogenicity of Magnaporthe oryzae
Erratum in
-
Correction: Eight RGS and RGS-like Proteins Orchestrate Growth, Differentiation, and Pathogenicity of Magnaporthe oryzae.PLoS Pathog. 2019 Nov 15;15(11):e1008187. doi: 10.1371/journal.ppat.1008187. eCollection 2019 Nov. PLoS Pathog. 2019. PMID: 31730634 Free PMC article.
Abstract
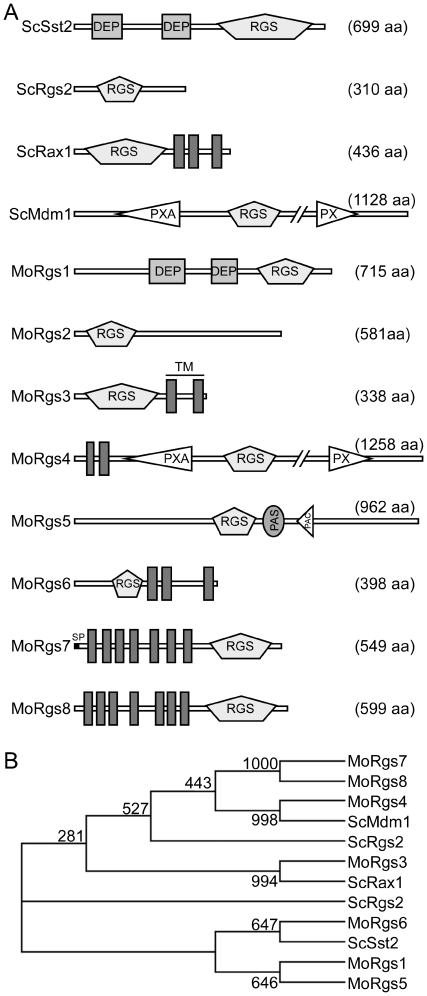
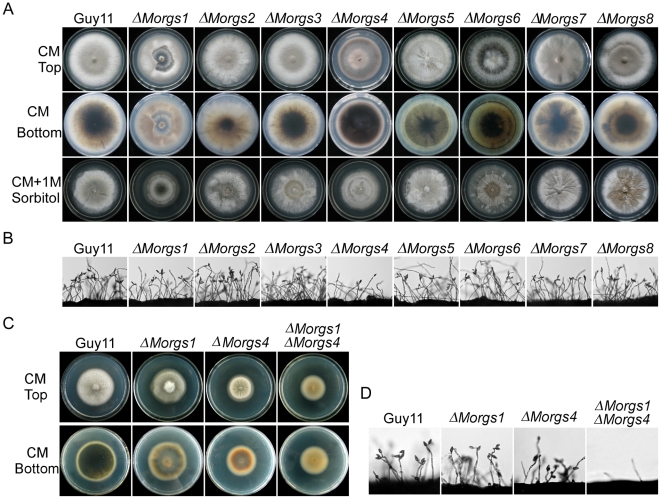
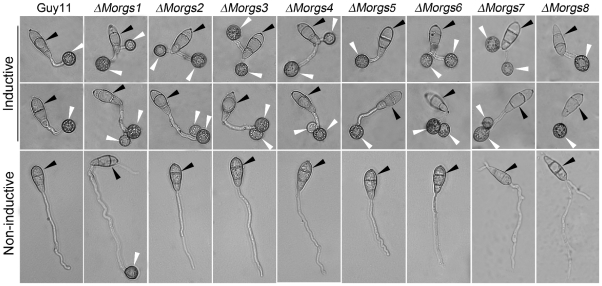
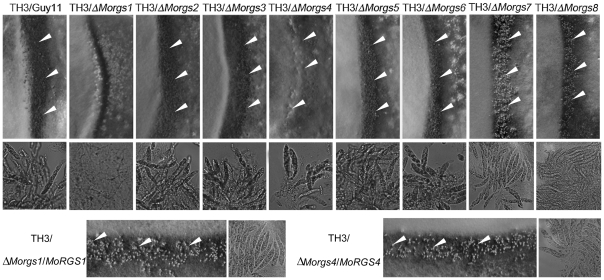
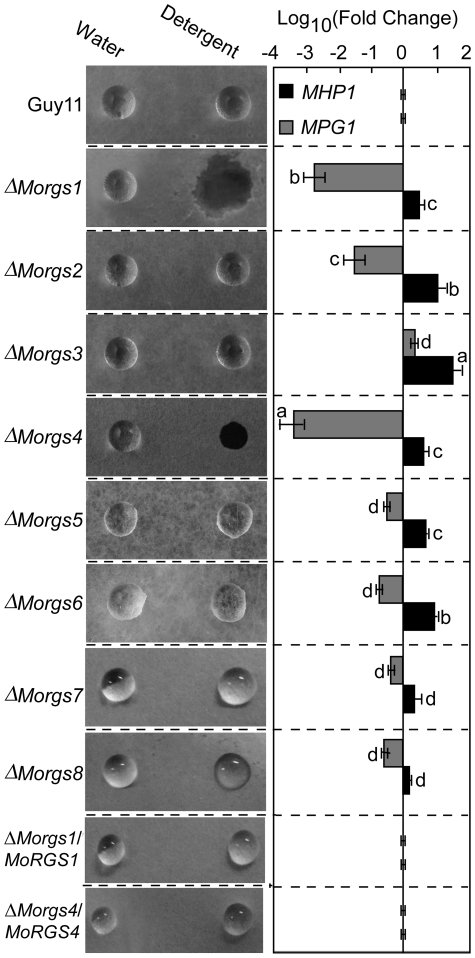
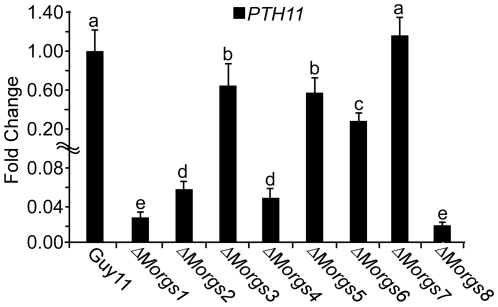
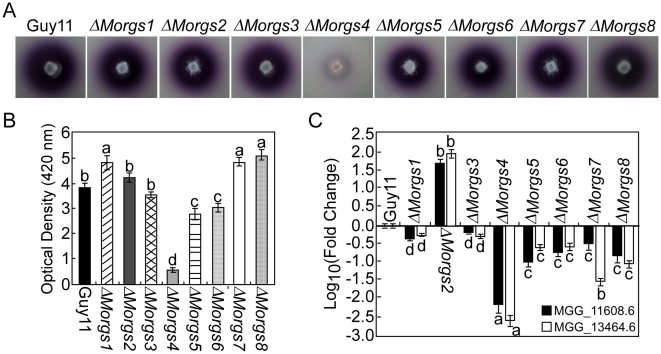
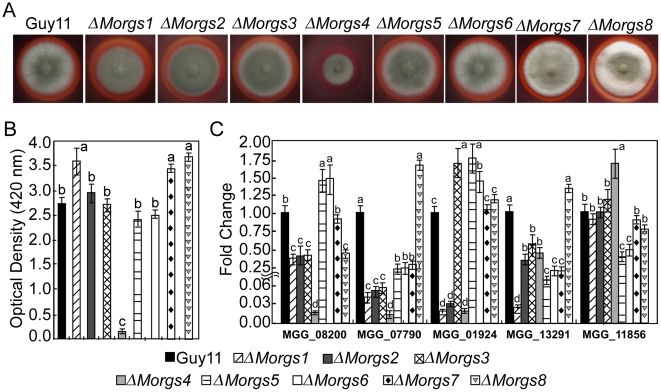
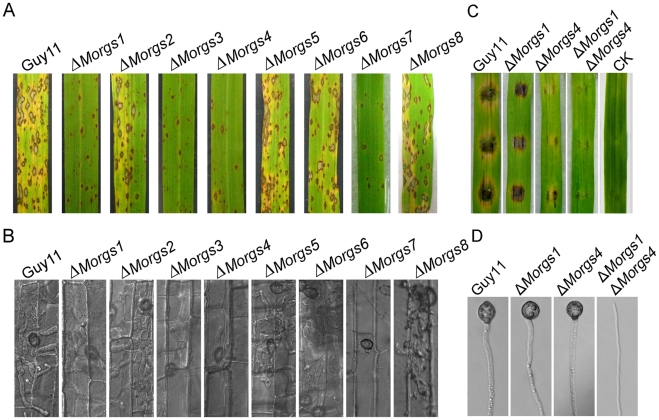
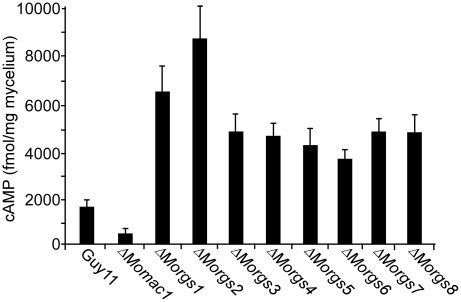
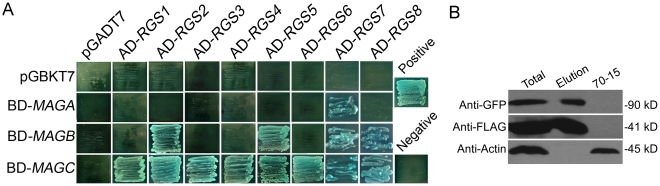
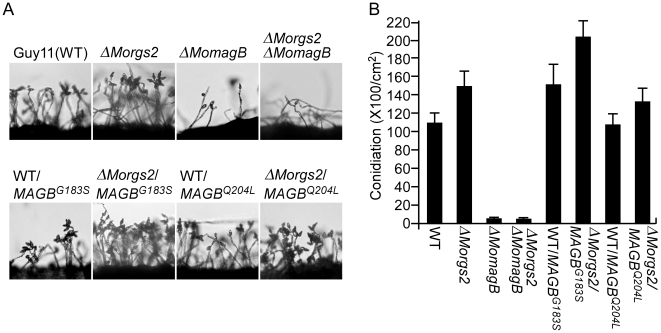
A previous study identified MoRgs1 as an RGS protein that negative regulates G-protein signaling to control developmental processes such as conidiation and appressorium formation in Magnaporthe oryzae. Here, we characterized additional seven RGS and RGS-like proteins (MoRgs2 through MoRgs8). We found that MoRgs1 and MoRgs4 positively regulate surface hydrophobicity, conidiation, and mating. Indifference to MoRgs1, MoRgs4 has a role in regulating laccase and peroxidase activities. MoRgs1, MoRgs2, MoRgs3, MoRgs4, MoRgs6, and MoRgs7 are important for germ tube growth and appressorium formation. Interestingly, MoRgs7 and MoRgs8 exhibit a unique domain structure in which the RGS domain is linked to a seven-transmembrane motif, a hallmark of G-protein coupled receptors (GPCRs). We have also shown that MoRgs1 regulates mating through negative regulation of Gα MoMagB and is involved in the maintenance of cell wall integrity. While all proteins appear to be involved in the control of intracellular cAMP levels, only MoRgs1, MoRgs3, MoRgs4, and MoRgs7 are required for full virulence. Taking together, in addition to MoRgs1 functions as a prominent RGS protein in M. oryzae, MoRgs4 and other RGS and RGS-like proteins are also involved in a complex process governing asexual/sexual development, appressorium formation, and pathogenicity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
The seven transmembrane domain protein MoRgs7 functions in surface perception and undergoes coronin MoCrn1-dependent endocytosis in complex with Gα subunit MoMagA to promote cAMP signaling and appressorium formation in Magnaporthe oryzae.PLoS Pathog. 2019 Feb 25;15(2):e1007382. doi: 10.1371/journal.ppat.1007382. eCollection 2019 Feb. PLoS Pathog. 2019. PMID: 30802274 Free PMC article.
-
Magnaporthe oryzae Auxiliary Activity Protein MoAa91 Functions as Chitin-Binding Protein To Induce Appressorium Formation on Artificial Inductive Surfaces and Suppress Plant Immunity.mBio. 2020 Mar 24;11(2):e03304-19. doi: 10.1128/mBio.03304-19. mBio. 2020. PMID: 32209696 Free PMC article.
-
MoRgs3 functions in intracellular reactive oxygen species perception-integrated cAMP signaling to promote appressorium formation in Magnaporthe oryzae.mBio. 2024 Aug 14;15(8):e0099624. doi: 10.1128/mbio.00996-24. Epub 2024 Jul 9. mBio. 2024. PMID: 38980036 Free PMC article.
-
MoMip11, a MoRgs7-interacting protein, functions as a scaffolding protein to regulate cAMP signaling and pathogenicity in the rice blast fungus Magnaporthe oryzae.Environ Microbiol. 2018 Sep;20(9):3168-3185. doi: 10.1111/1462-2920.14102. Epub 2018 May 15. Environ Microbiol. 2018. PMID: 29727050 Free PMC article.
-
Characterizations and functions of regulator of G protein signaling (RGS) in fungi.Appl Microbiol Biotechnol. 2013 Sep;97(18):7977-87. doi: 10.1007/s00253-013-5133-1. Epub 2013 Aug 6. Appl Microbiol Biotechnol. 2013. PMID: 23917634 Review.
Cited by
-
Advancement in the Breeding, Biotechnological and Genomic Tools towards Development of Durable Genetic Resistance against the Rice Blast Disease.Plants (Basel). 2022 Sep 13;11(18):2386. doi: 10.3390/plants11182386. Plants (Basel). 2022. PMID: 36145787 Free PMC article. Review.
-
MoImd4 mediates crosstalk between MoPdeH-cAMP signalling and purine metabolism to govern growth and pathogenicity in Magnaporthe oryzae.Mol Plant Pathol. 2019 Apr;20(4):500-518. doi: 10.1111/mpp.12770. Epub 2019 Jan 11. Mol Plant Pathol. 2019. PMID: 30426699 Free PMC article.
-
MoVrp1, a putative verprolin protein, is required for asexual development and infection in the rice blast fungus Magnaporthe oryzae.Sci Rep. 2017 Jan 24;7:41148. doi: 10.1038/srep41148. Sci Rep. 2017. PMID: 28117435 Free PMC article.
-
Functional and topological diversity of LOV domain photoreceptors.Proc Natl Acad Sci U S A. 2016 Mar 15;113(11):E1442-51. doi: 10.1073/pnas.1509428113. Epub 2016 Feb 29. Proc Natl Acad Sci U S A. 2016. PMID: 26929367 Free PMC article.
-
MoErv14 mediates the intracellular transport of cell membrane receptors to govern the appressorial formation and pathogenicity of Magnaporthe oryzae.PLoS Pathog. 2023 Apr 3;19(4):e1011251. doi: 10.1371/journal.ppat.1011251. eCollection 2023 Apr. PLoS Pathog. 2023. PMID: 37011084 Free PMC article.
References
-
- Malbon CC. G proteins in development. Nat Rev Mol Cell Bio. 2005;6:689–701. - PubMed
-
- Dohlman HG, Thorner JW. Regulation of G protein-initiated signal transduction in yeast: Paradigms and principles. Annu Rev Biochem. 2001;70:703–754. - PubMed
-
- Clapham DE, Neer EJ. G protein beta gamma subunits. Annu Rev Pharmacol Toxicol. 1997;37:167–203. - PubMed
-
- Hamm HE. The many faces of G protein signaling. J Biol Chem. 1998;273:669–672. - PubMed
-
- Neer EJ. Heterotrimeric G-proteins - organizers of transmembrane signals. Cell. 1995;80:249–257. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
