

Substitutions in the amino-terminal tail of neurospora histone H3 have varied effects on DNA methylation
- PMID: 22242002
- PMCID: PMC3248561
- DOI: 10.1371/journal.pgen.1002423
Substitutions in the amino-terminal tail of neurospora histone H3 have varied effects on DNA methylation
Abstract
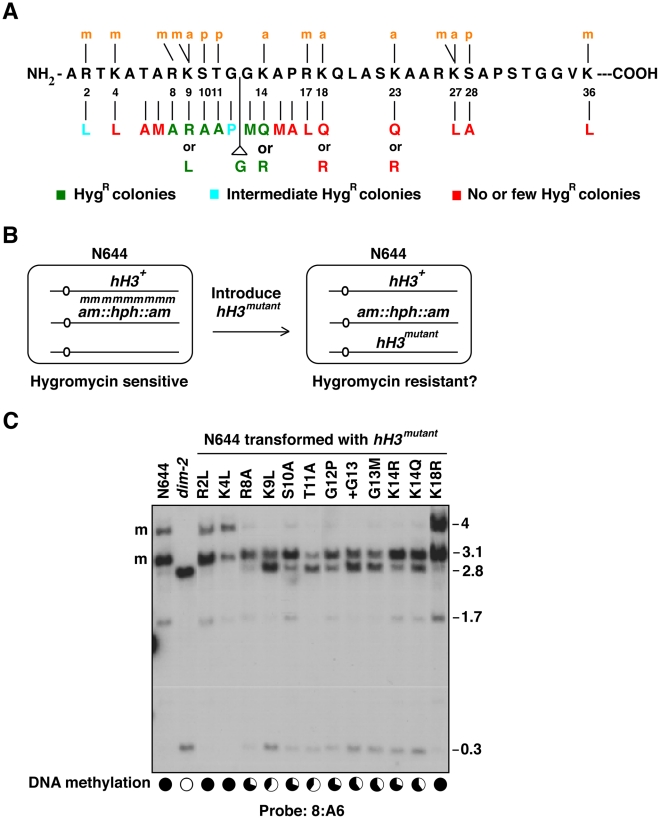
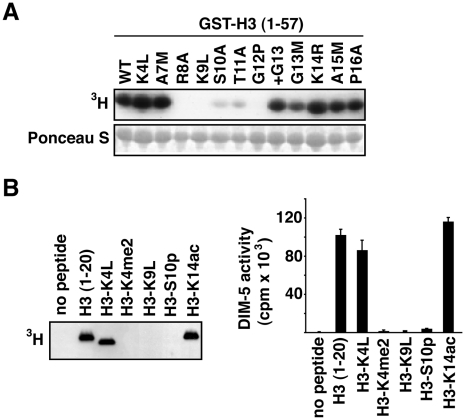
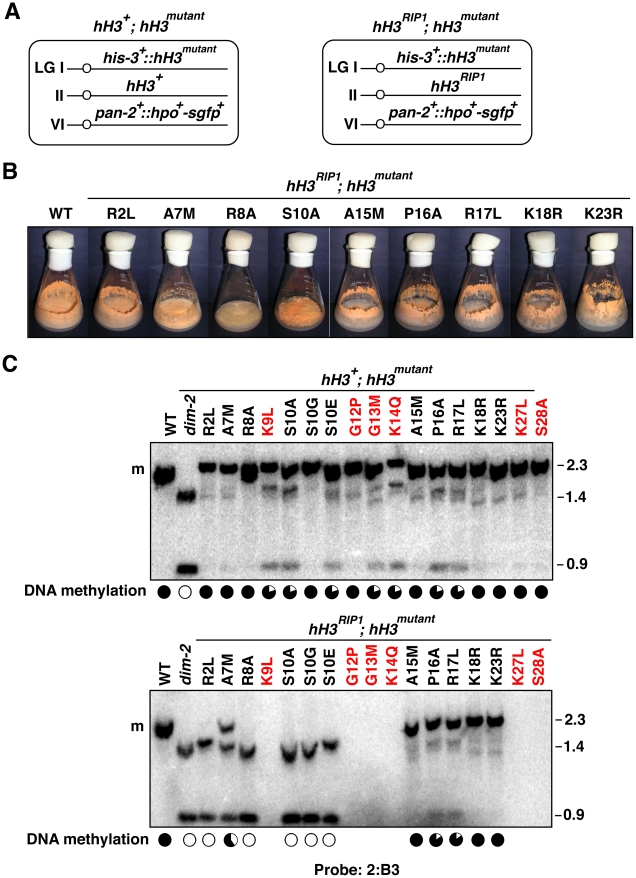
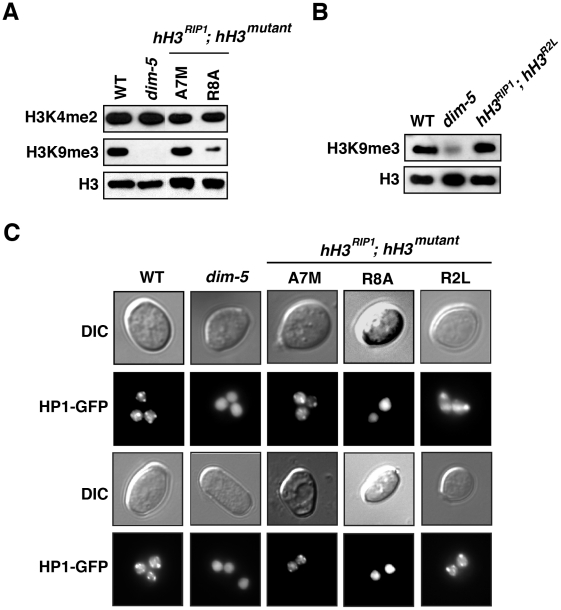
Eukaryotic genomes are partitioned into active and inactive domains called euchromatin and heterochromatin, respectively. In Neurospora crassa, heterochromatin formation requires methylation of histone H3 at lysine 9 (H3K9) by the SET domain protein DIM-5. Heterochromatin protein 1 (HP1) reads this mark and directly recruits the DNA methyltransferase, DIM-2. An ectopic H3 gene carrying a substitution at K9 (hH3(K9L) or hH3(K9R)) causes global loss of DNA methylation in the presence of wild-type hH3 (hH3(WT)). We investigated whether other residues in the N-terminal tail of H3 are important for methylation of DNA and of H3K9. Mutations in the N-terminal tail of H3 were generated and tested for effects in vitro and in vivo, in the presence or absence of the wild-type allele. Substitutions at K4, K9, T11, G12, G13, K14, K27, S28, and K36 were lethal in the absence of a wild-type allele. In contrast, mutants bearing substitutions of R2, A7, R8, S10, A15, P16, R17, K18, and K23 were viable. The effect of substitutions on DNA methylation were variable; some were recessive and others caused a semi-dominant loss of DNA methylation. Substitutions of R2, A7, R8, S10, T11, G12, G13, K14, and P16 caused partial or complete loss of DNA methylation in vivo. Only residues R8-G12 were required for DIM-5 activity in vitro. DIM-5 activity was inhibited by dimethylation of H3K4 and by phosphorylation of H3S10, but not by acetylation of H3K14. We conclude that the H3 tail acts as an integrating platform for signals that influence DNA methylation, in part through methylation of H3K9.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
