

Precursor uptake assays and metabolic analyses in isolated tomato fruit chromoplasts
- PMID: 22243738
- PMCID: PMC3269359
- DOI: 10.1186/1746-4811-8-1
Precursor uptake assays and metabolic analyses in isolated tomato fruit chromoplasts
Abstract
Background: Carotenoids are the most widespread group of pigments found in nature. In addition to their role in the physiology of the plant, carotenoids also have nutritional relevance as their incorporation in the human diet provides health benefits. In non-photosynthetic tissues, carotenoids are synthesized and stored in specialized plastids called chromoplasts. At present very little is known about the origin of the metabolic precursors and cofactors required to sustain the high rate of carotenoid biosynthesis in these plastids. Recent proteomic data have revealed a number of biochemical and metabolic processes potentially operating in fruit chromoplasts. However, considering that chloroplast to chromoplast differentiation is a very rapid process during fruit ripening, there is the possibility that some of the proteins identified in the proteomic analysis could represent remnants no longer having a functional role in chromoplasts. Therefore, experimental validation is necessary to prove whether these predicted processes are actually operative in chromoplasts.
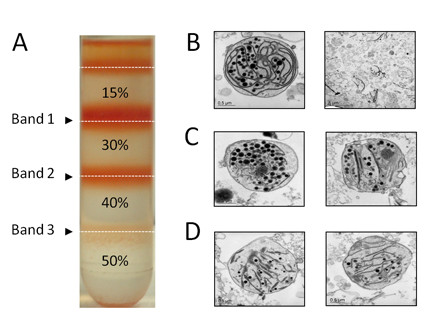
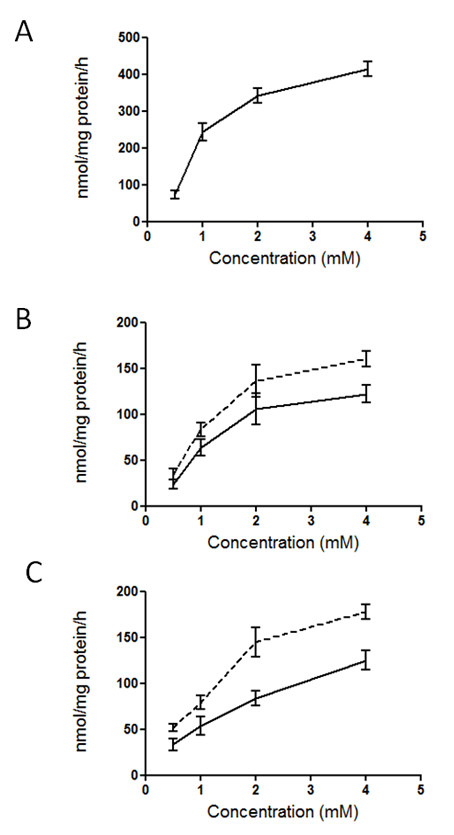
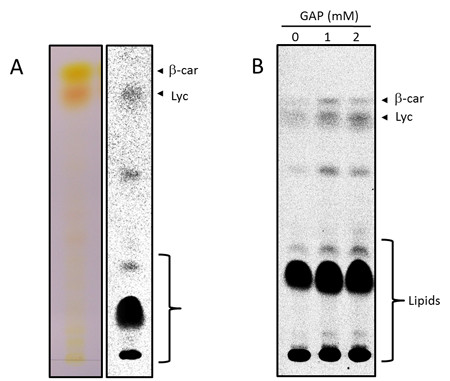
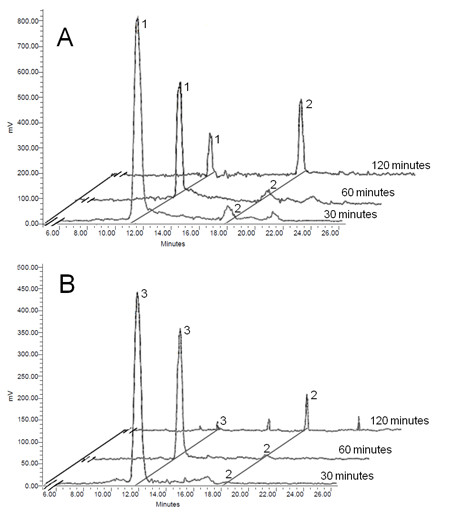
Results: A method has been established for high-yield purification of tomato fruit chromoplasts suitable for metabolic studies. Radiolabeled precursors were efficiently incorporated and further metabolized in isolated chromoplast. Analysis of labeled lipophilic compounds has revealed that lipid biosynthesis is a very efficient process in chromoplasts, while the relatively low incorporation levels found in carotenoids suggest that lipid production may represent a competing pathway for carotenoid biosynthesis. Malate and pyruvate are efficiently converted into acetyl-CoA, in agreement with the active operation of the malic enzyme and the pyruvate dehydrogenase complex in the chromoplast. Our results have also shown that isolated chromoplasts can actively sustain anabolic processes without the exogenous supply of ATP, thus suggesting that these organelles may generate this energetic cofactor in an autonomous way.
Conclusions: We have set up a method for high yield purification of intact tomato fruit chromoplasts suitable for precursor uptake assays and metabolic analyses. Using targeted radiolabeled precursors we have been able to unravel novel biochemical and metabolic aspects related with carotenoid and lipid biosynthesis in tomato fruit chromoplasts. The reported chromoplast system could represent a valuable platform to address the validation and characterization of functional processes predicted from recent transcriptomic and proteomic data.
Figures
References
-
- Beyer P, Al-Babili S, Ye X, Lucca P, Schaub P, Welsch R, Potrykus I. Golden Rice: Introducing the β-Carotene Biosynthesis Pathway into Rice Endosperm by Genetic Engineering to Defeat Vitamin A Deficiency. J Nutr. 2002;132:506S–510S. - PubMed
-
- Camara B, Hugueney P, Bouvier F, Kuntz M, Monéger R. Biochemistry and molecular biology of chromoplast development. Int Rev Cytol. 1995;163:175–247. - PubMed
