

Extrasynaptic GABA(A) receptors: their function in the CNS and implications for disease
- PMID: 22243744
- PMCID: PMC3399243
- DOI: 10.1016/j.neuron.2011.12.012
Extrasynaptic GABA(A) receptors: their function in the CNS and implications for disease
Abstract
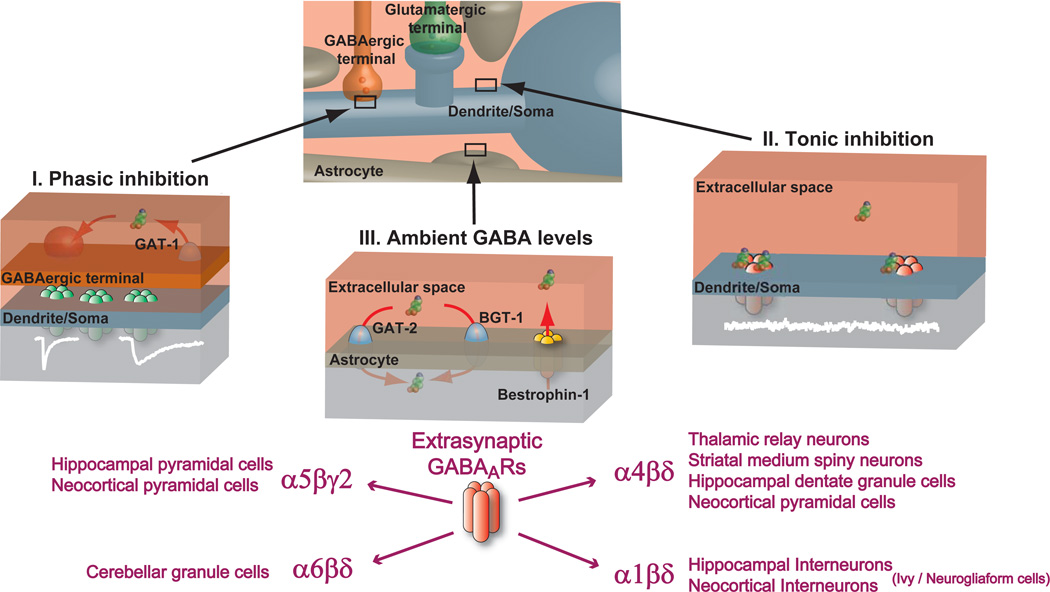
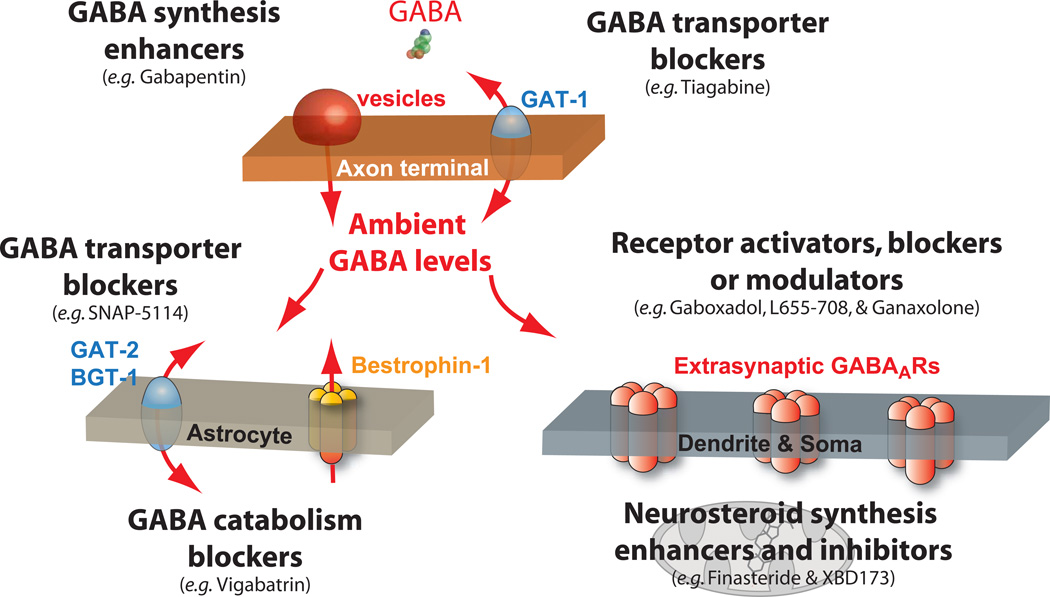
Over the past two decades, research has identified extrasynaptic GABA(A) receptor populations that enable neurons to sense the low ambient GABA concentrations present in the extracellular space in order to generate a form of tonic inhibition not previously considered in studies of neuronal excitability. The importance of this tonic inhibition in regulating states of consciousness is highlighted by the fact that extrasynaptic GABA(A) receptors (GABA(A)Rs) are believed to be key targets for anesthetics, sleep-promoting drugs, neurosteroids, and alcohol. The neurosteroid sensitivity of these extrasynaptic GABA(A)Rs may explain their importance in stress-, ovarian cycle-, and pregnancy-related mood disorders. Moreover, disruptions in network dynamics associated with schizophrenia, epilepsy, and Parkinson's disease may well involve alterations in the tonic GABA(A)R-mediated conductance. Extrasynaptic GABA(A)Rs may therefore present a therapeutic target for treatment of these diseases, with the potential to enhance cognition and aid poststroke functional recovery.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Agnati LF, Guidolin D, Guescini M, Genedani S, Fuxe K. Understanding wiring and volume transmission. Brain Res Rev. 2010;64:137–159. - PubMed
-
- Atack JR. Preclinical and clinical pharmacology of the GABAA receptor alpha5 subtype-selective inverse agonist alpha5IA. Pharmacol Ther. 2010;125:11–26. - PubMed
-
- Bai D, Zhu G, Pennefather P, Jackson MF, MacDonald JF, Orser BA. Distinct functional and pharmacological properties of tonic and quantal inhibitory postsynaptic currents mediated by gamma-aminobutyric acid(A) receptors in hippocampal neurons. Mol Pharmacol. 2001;59:814–824. - PubMed
-
- Ballard TM, Knoflach F, Prinssen E, Borroni E, Vivian JA, Basile J, Gasser R, Moreau JL, Wettstein JG, Buettelmann B, et al. RO4938581, a novel cognitive enhancer acting at GABAA alpha5 subunit-containing receptors. Psychopharmacology (Berl) 2009;202:207–223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
