

Otic mesenchyme cells regulate spiral ganglion axon fasciculation through a Pou3f4/EphA4 signaling pathway
- PMID: 22243746
- PMCID: PMC3259535
- DOI: 10.1016/j.neuron.2011.10.029
Otic mesenchyme cells regulate spiral ganglion axon fasciculation through a Pou3f4/EphA4 signaling pathway
Abstract
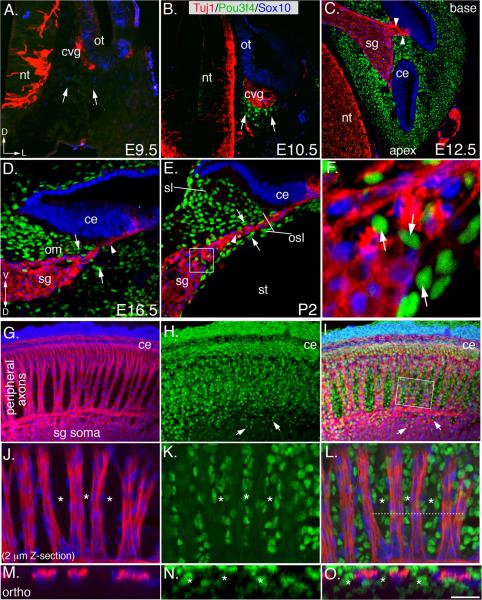
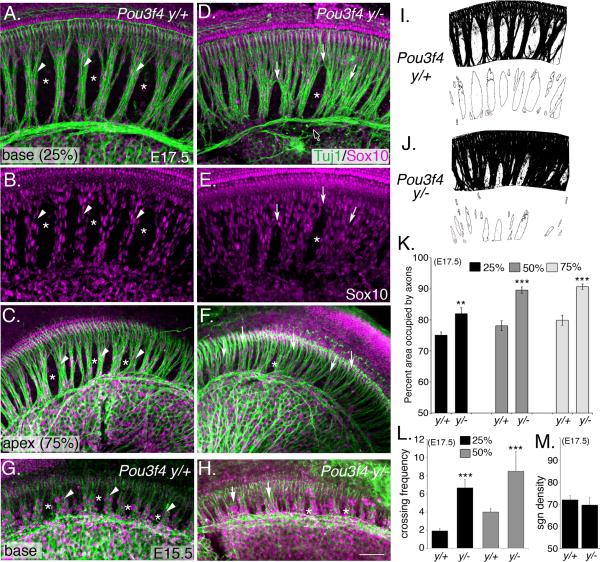
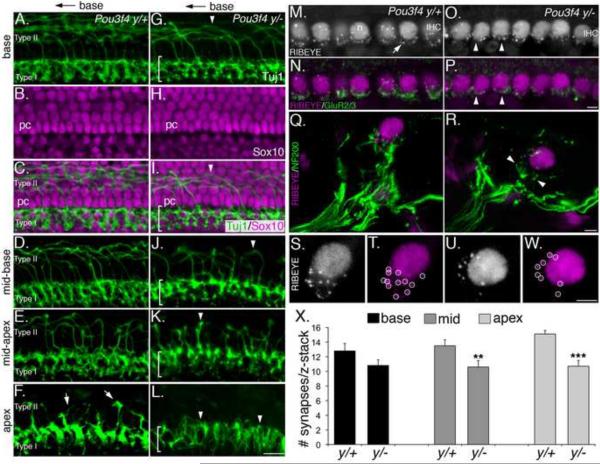
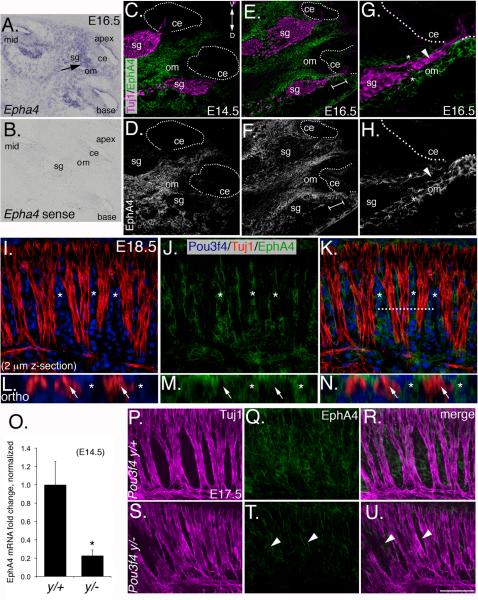
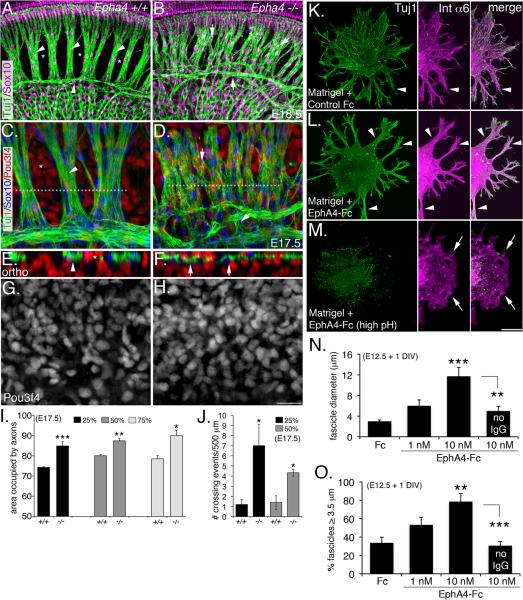
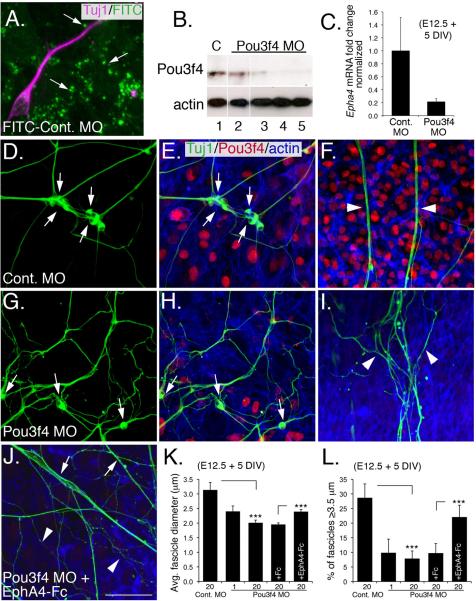
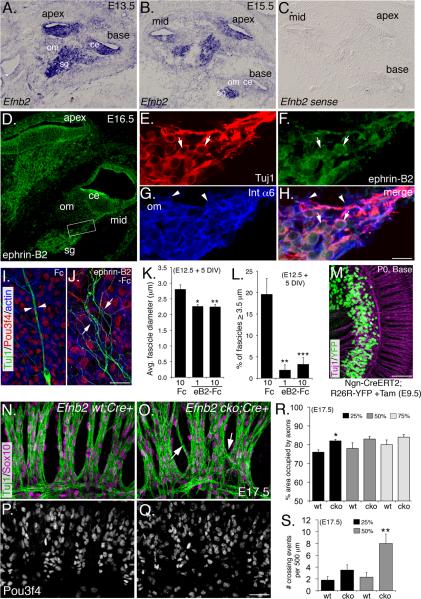
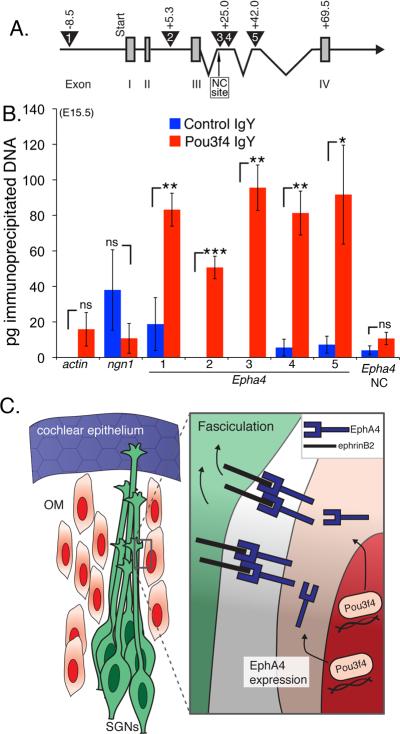
Peripheral axons from auditory spiral ganglion neurons (SGNs) form an elaborate series of radially and spirally oriented projections that interpret complex aspects of the auditory environment. However, the developmental processes that shape these axon tracts are largely unknown. Radial bundles are comprised of dense SGN fascicles that project through otic mesenchyme to form synapses within the cochlea. Here, we show that radial bundle fasciculation and synapse formation are disrupted when Pou3f4 (DFNX2) is deleted from otic mesenchyme. Further, we demonstrate that Pou3f4 binds to and directly regulates expression of Epha4, Epha4⁻/⁻ mice present similar SGN defects, and exogenous EphA4 promotes SGN fasciculation in the absence of Pou3f4. Finally, Efnb2 deletion in SGNs leads to similar fasciculation defects, suggesting that ephrin-B2/EphA4 interactions are critical during this process. These results indicate a model whereby Pou3f4 in the otic mesenchyme establishes an Eph/ephrin-mediated fasciculation signal that promotes inner radial bundle formation.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Ahn KJ, Passero F, Jr., Crenshaw EB., 3rd Otic mesenchyme expression of Cre recombinase directed by the inner ear enhancer of the Brn4/Pou3f4 gene. Genesis. 2009;47:137–141. - PubMed
-
- Baum PD, Garriga G. Neuronal migrations and axon fasciculation are disrupted in ina-1 integrin mutants. Neuron. 1997;19:51–62. - PubMed
-
- Bianchi LM, Gale NW. Distribution of Eph-related molecules in the developing and mature cochlea. Hear Res. 1998;117:161–172. - PubMed
-
- Bossing T, Brand AH. Dephrin, a transmembrane ephrin with a unique structure, prevents interneuronal axons from exiting the Drosophila embryonic CNS. Development. 2002;129:4205–4218. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
