

Reciprocal inhibition of inhibition: a circuit motif for flexible categorization in stimulus selection
- PMID: 22243757
- PMCID: PMC3263974
- DOI: 10.1016/j.neuron.2011.10.037
Reciprocal inhibition of inhibition: a circuit motif for flexible categorization in stimulus selection
Abstract
As a precursor to the selection of a stimulus for gaze and attention, a midbrain network categorizes stimuli into "strongest" and "others." The categorization tracks flexibly, in real time, the absolute strength of the strongest stimulus. In this study, we take a first-principles approach to computations that are essential for such categorization. We demonstrate that classical feedforward lateral inhibition cannot produce flexible categorization. However, circuits in which the strength of lateral inhibition varies with the relative strength of competing stimuli categorize successfully. One particular implementation--reciprocal inhibition of feedforward lateral inhibition--is structurally the simplest, and it outperforms others in flexibly categorizing rapidly and reliably. Strong predictions of this anatomically supported circuit model are validated by neural responses measured in the owl midbrain. The results demonstrate the extraordinary power of a remarkably simple, neurally grounded circuit motif in producing flexible categorization, a computation fundamental to attention, perception, and decision making.
Copyright © 2012 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures



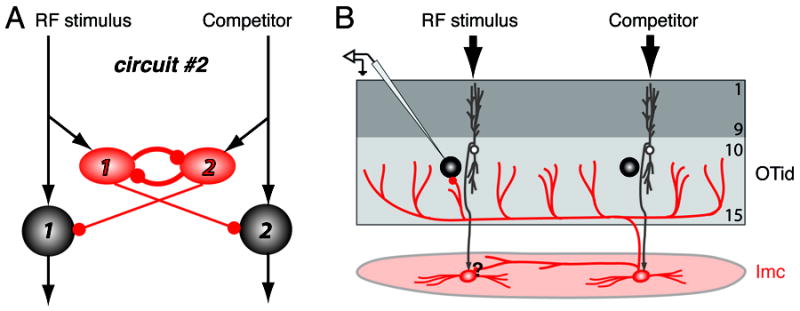

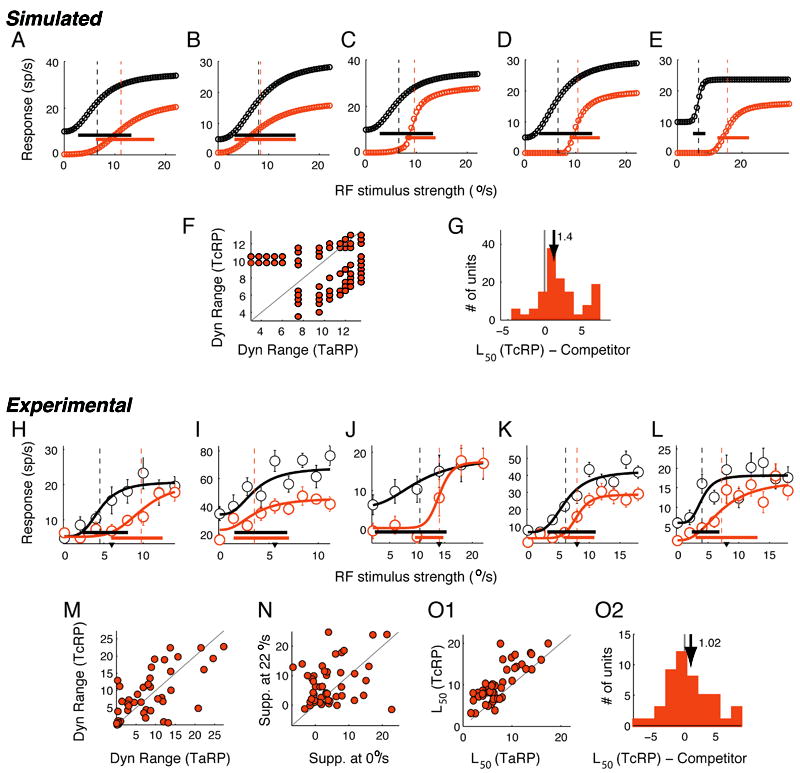

Comment in
-
Adaptive switches in midbrain circuits.Neuron. 2012 Jan 12;73(1):6-7. doi: 10.1016/j.neuron.2011.12.017. Neuron. 2012. PMID: 22243742 Free PMC article.
References
-
- Allman J, Miezin F, McGuinness E. Stimulus specific responses from beyond the classical receptive field: neurophysiological mechanisms for local-global comparisons in visual neurons. Annu Rev Neurosci. 1985;8:407–430. - PubMed
-
- Ayaz A, Chance FS. Gain modulation of neuronal responses by subtractive and divisive mechanisms of inhibition. J Neurophysiol. 2009;101:958–968. - PubMed
-
- Blakemore C, Tobin EA. Lateral inhibition between orientation detectors in the cat’s visual cortex. Exp Brain Res. 1972;15:439–440. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
