

Double strand binding-single strand incision mechanism for human flap endonuclease: implications for the superfamily
- PMID: 22244820
- PMCID: PMC3345168
- DOI: 10.1016/j.mad.2011.11.009
Double strand binding-single strand incision mechanism for human flap endonuclease: implications for the superfamily
Abstract
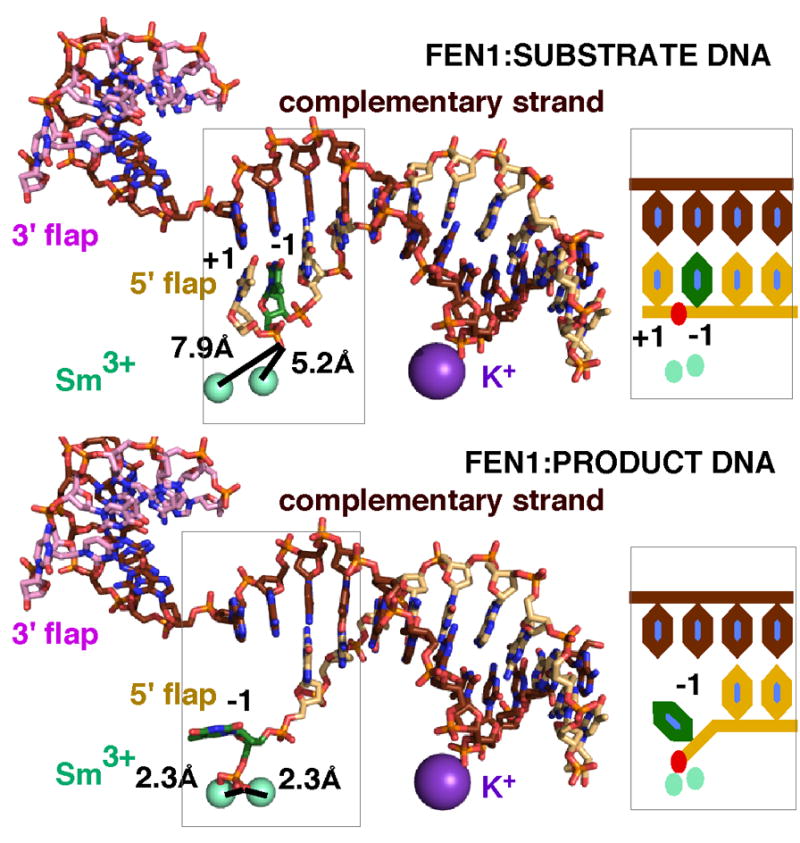
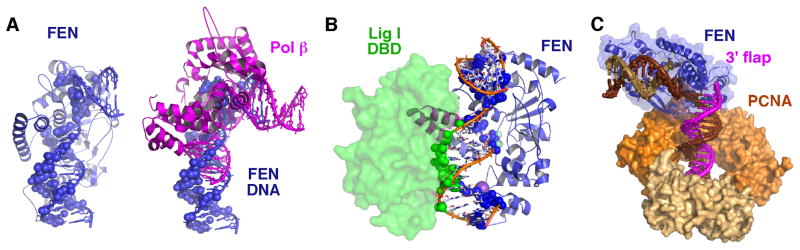
Detailed structural, mutational, and biochemical analyses of human FEN1/DNA complexes have revealed the mechanism for recognition of 5' flaps formed during lagging strand replication and DNA repair. FEN1 processes 5' flaps through a previously unknown, but structurally elegant double-stranded (ds) recognition/single stranded (ss) incision mechanism that both selects for 5' flaps and selects against ss DNA or RNA, intact dsDNA, and 3' flaps. Two major DNA binding interfaces, including a K(+) bridge between the DNA and the H2TH motif, are spaced one helical turn apart and together select for substrates with dsDNA. A conserved helical gateway and a helical cap protects the two-metal active site and selects for ss flaps with free termini. Structures of substrate and product reveal an unusual step between binding substrate and incision that involves a double base unpairing with incision occurring in the resulting unpaired DNA or RNA. Ordering of the active site requires a disorder-to-order transition induced by binding of an unpaired 3' flap, which ensures that the product is ligatable. Comparison with FEN superfamily members, including XPG, EXO1, and GEN1, identifies superfamily motifs such as the helical gateway that select for ss-dsDNA junctions and provides key biological insights into nuclease specificity and regulation.
Copyright © 2011 Elsevier Ireland Ltd. All rights reserved.
Figures



References
-
- Bornarth CJ, Ranalli TA, Henricksen LA, Wahl AF, Bambara RA. Effect of flap modifications on human FEN1 cleavage. Biochemistry. 1999;38:13347–13354. - PubMed
-
- Ceska TA, Sayers JR, Stier G, Suck D. A helical arch allowing singlestranded DNA to thread through T5 5′-exonuclease. Nature. 1996;382:90–93. - PubMed
-
- Chapados BR, Hosfield DJ, Han S, Qiu J, Yelent B, Shen B, Tainer JA. Structural basis for FEN-1 substrate specificity and PCNA-mediated activation in DNA replication and repair. Cell. 2004;116:39–50. - PubMed
-
- Cvetkovic A, Menon AL, Thorgersen MP, Scott JW, Poole FL, 2nd, Jenney FE, Jr, Lancaster WA, Praissman JL, Shanmukh S, Vaccaro BJ, Trauger SA, Kalisiak E, Apon JV, Siuzdak G, Yannone SM, Tainer JA, Adams MW. Microbial metalloproteomes are largely uncharacterized. Nature. 2010;466:779–782. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
