

Glucose-6-phosphatase-β, implicated in a congenital neutropenia syndrome, is essential for macrophage energy homeostasis and functionality
- PMID: 22246029
- PMCID: PMC3350367
- DOI: 10.1182/blood-2011-09-377820
Glucose-6-phosphatase-β, implicated in a congenital neutropenia syndrome, is essential for macrophage energy homeostasis and functionality
Abstract
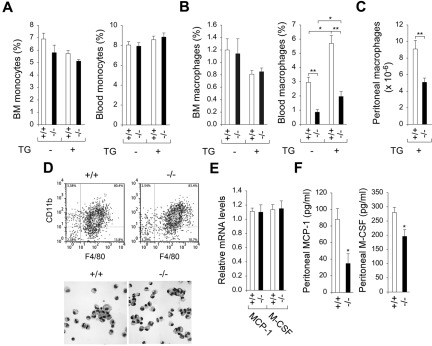
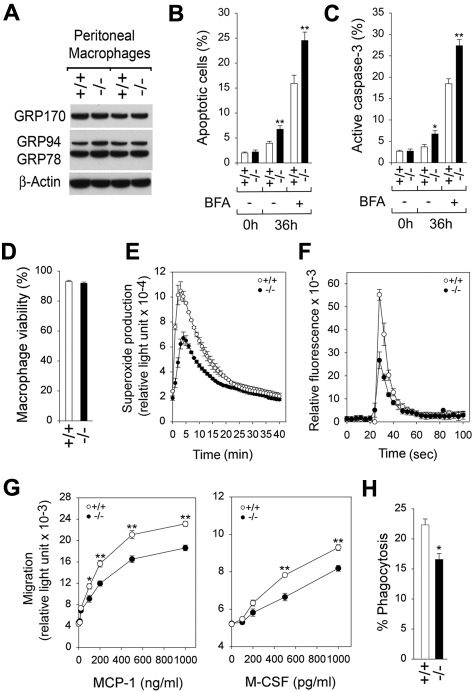
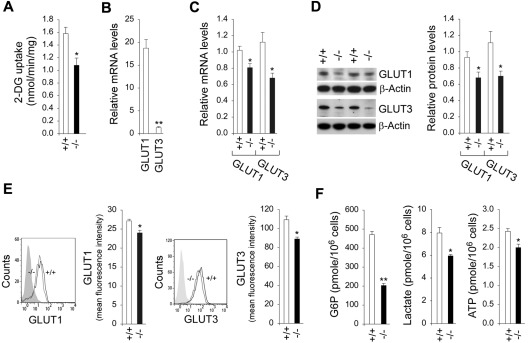
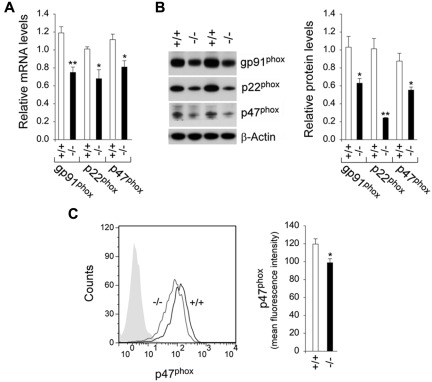
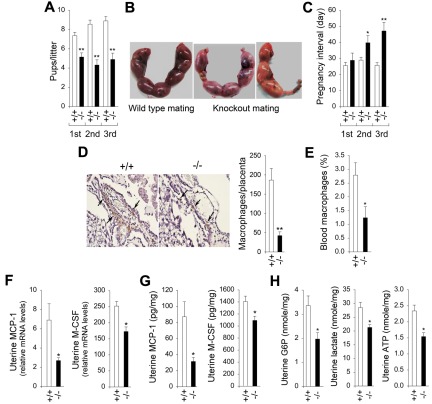
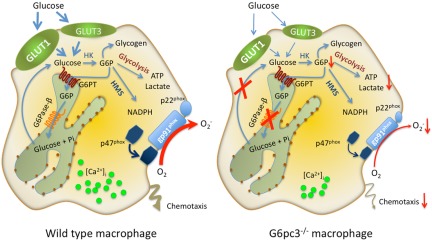
Glucose-6-phosphatase-β (G6Pase-β or G6PC3) deficiency, also known as severe congenital neutropenia syndrome 4, is characterized not only by neutropenia but also by impaired neutrophil energy homeostasis and functionality. We now show the syndrome is also associated with macrophage dysfunction, with murine G6pc3(-/-) macrophages having impairments in their respiratory burst, chemotaxis, calcium flux, and phagocytic activities. Consistent with a glucose-6-phosphate (G6P) metabolism deficiency, G6pc3(-/-) macrophages also have a lower glucose uptake and lower levels of G6P, lactate, and ATP than wild-type macrophages. Furthermore, the expression of NADPH oxidase subunits and membrane translocation of p47(phox) are down-regulated, and G6pc3(-/-) macrophages exhibit repressed trafficking in vivo both during an inflammatory response and in pregnancy. During pregnancy, the absence of G6Pase-β activity also leads to impaired energy homeostasis in the uterus and reduced fertility of G6pc3(-/-) mothers. Together these results show that immune deficiencies in this congenital neutropenia syndrome extend beyond neutrophil dysfunction.
Figures
References
-
- Shieh J-J, Pan C-J, Mansfield BC, Chou JY. Glucose-6-phosphate hydrolase, widely expressed outside the liver, can explain age-dependent resolution of hypoglycemia in glycogen storage disease type Ia. J Biol Chem. 2003;278(47):47098–47103. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
