

GLUT4 exocytosis
- PMID: 22247191
- PMCID: PMC3258103
- DOI: 10.1242/jcs.097063
GLUT4 exocytosis
Abstract
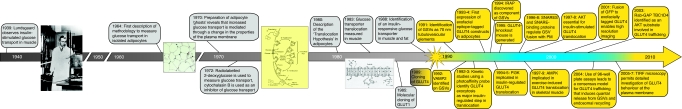
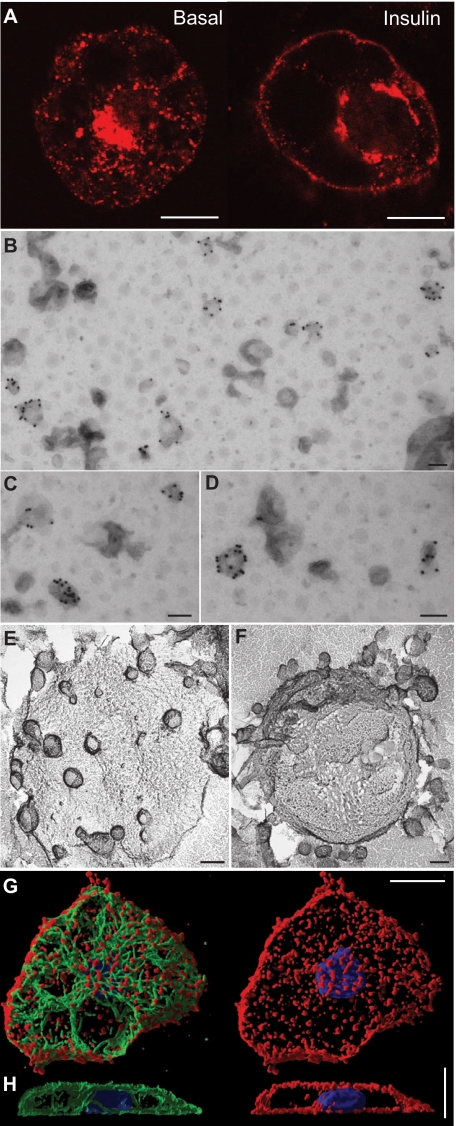
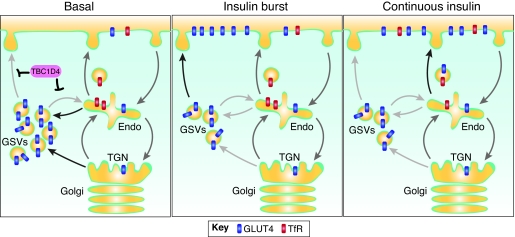
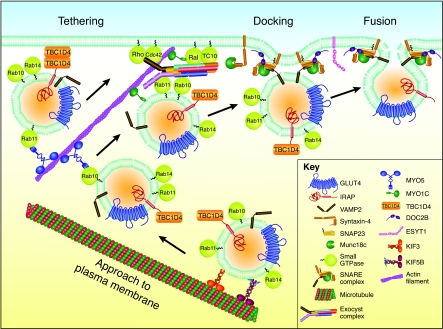
GLUT4 is an insulin-regulated glucose transporter that is responsible for insulin-regulated glucose uptake into fat and muscle cells. In the absence of insulin, GLUT4 is mainly found in intracellular vesicles referred to as GLUT4 storage vesicles (GSVs). Here, we summarise evidence for the existence of these specific vesicles, how they are sequestered inside the cell and how they undergo exocytosis in the presence of insulin. In response to insulin stimulation, GSVs fuse with the plasma membrane in a rapid burst and in the continued presence of insulin GLUT4 molecules are internalised and recycled back to the plasma membrane in vesicles that are distinct from GSVs and probably of endosomal origin. In this Commentary we discuss evidence that this delivery process is tightly regulated and involves numerous molecules. Key components include the actin cytoskeleton, myosin motors, several Rab GTPases, the exocyst, SNARE proteins and SNARE regulators. Each step in this process is carefully orchestrated in a sequential and coupled manner and we are beginning to dissect key nodes within this network that determine vesicle-membrane fusion in response to insulin. This regulatory process clearly involves the Ser/Thr kinase AKT and the exquisite manner in which this single metabolic process is regulated makes it a likely target for lesions that might contribute to metabolic disease.
Figures
References
-
- Abel E. D., Peroni O., Kim J. K., Kim Y. B., Boss O., Hadro E., Minnemann T., Shulman G. I., Kahn B. B. (2001). Adipose-selective targeting of the GLUT4 gene impairs insulin action in muscle and liver. Nature 409, 729-733 - PubMed
-
- Aledo J. C., Darakhshan F., Hundal H. S. (1995). Rab4, but not the transferrin receptor, is colocalized with GLUT4 in an insulin-sensitive intracellular compartment in rat skeletal muscle. Biochem. Biophys. Res. Commun. 215, 321-328 - PubMed
-
- Antonescu C. N., Diaz M., Femia G., Planas J. V., Klip A. (2008). Clathrin-dependent and independent endocytosis of glucose transporter 4 (GLUT4) in myoblasts: regulation by mitochondrial uncoupling. Traffic 9, 1173-1190 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
