

The epigenetic H3S10 phosphorylation mark is required for counteracting heterochromatic spreading and gene silencing in Drosophila melanogaster
- PMID: 22247192
- PMCID: PMC3258113
- DOI: 10.1242/jcs.092585
The epigenetic H3S10 phosphorylation mark is required for counteracting heterochromatic spreading and gene silencing in Drosophila melanogaster
Abstract
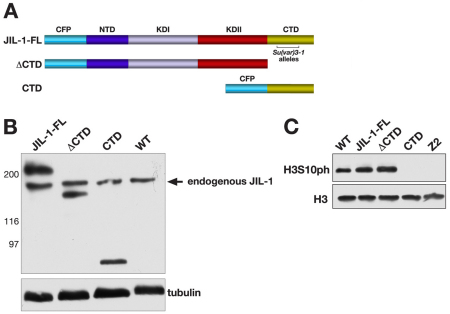
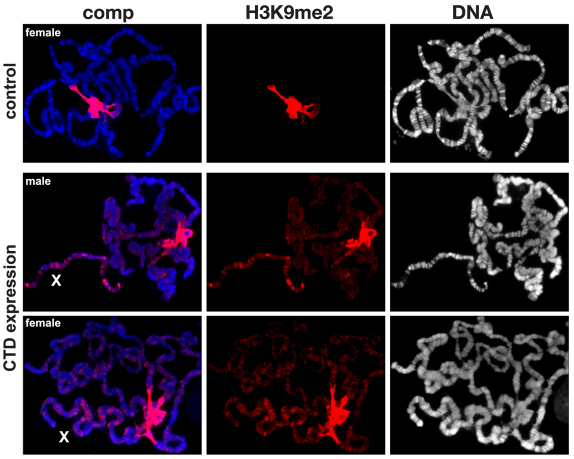
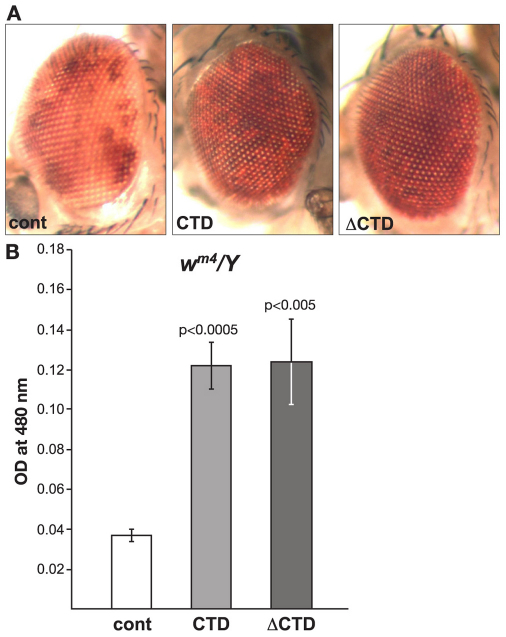
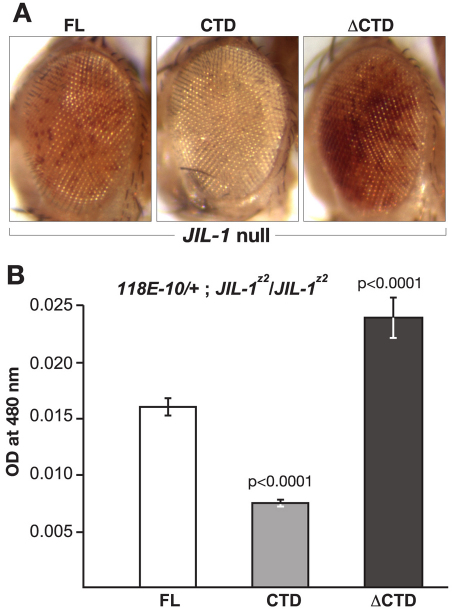
The JIL-1 kinase localizes specifically to euchromatin interband regions of polytene chromosomes and is the kinase responsible for histone H3S10 phosphorylation at interphase. Genetic interaction assays with strong JIL-1 hypomorphic loss-of-function alleles have demonstrated that the JIL-1 protein can counterbalance the effect of the major heterochromatin components on position-effect variegation (PEV) and gene silencing. However, it is unclear whether this was a causative effect of the epigenetic H3S10 phosphorylation mark, or whether the effect of the JIL-1 protein on PEV was in fact caused by other functions or structural features of the protein. By transgenically expressing various truncated versions of JIL-1, with or without kinase activity, and assessing their effect on PEV and heterochromatic spreading, we show that the gross perturbation of polytene chromosome morphology observed in JIL-1 null mutants is unrelated to gene silencing in PEV and is likely to occur as a result of faulty polytene chromosome alignment and/or organization, separate from epigenetic regulation of chromatin structure. Furthermore, the findings provide evidence that the epigenetic H3S10 phosphorylation mark itself is necessary for preventing the observed heterochromatic spreading independently of any structural contributions from the JIL-1 protein.
Figures

Similar articles
-
Domain requirements of the JIL-1 tandem kinase for histone H3 serine 10 phosphorylation and chromatin remodeling in vivo.J Biol Chem. 2013 Jul 5;288(27):19441-9. doi: 10.1074/jbc.M113.464271. Epub 2013 May 30. J Biol Chem. 2013. PMID: 23723094 Free PMC article.
-
H3S10 phosphorylation by the JIL-1 kinase regulates H3K9 dimethylation and gene expression at the white locus in Drosophila.Fly (Austin). 2012 Apr-Jun;6(2):93-7. doi: 10.4161/fly.20029. Epub 2012 Apr 1. Fly (Austin). 2012. PMID: 22634714 Free PMC article.
-
Histone H3S10 phosphorylation by the JIL-1 kinase in pericentric heterochromatin and on the fourth chromosome creates a composite H3S10phK9me2 epigenetic mark.Chromosoma. 2014 Jun;123(3):273-80. doi: 10.1007/s00412-014-0450-4. Epub 2014 Jan 16. Chromosoma. 2014. PMID: 24429699 Free PMC article.
-
Histone modification and the control of heterochromatic gene silencing in Drosophila.Chromosome Res. 2006;14(4):377-92. doi: 10.1007/s10577-006-1066-1. Chromosome Res. 2006. PMID: 16821134 Review.
-
Loss-of-function alleles of the JIL-1 kinase are strong suppressors of position effect variegation of the wm4 allele in Drosophila.Genetics. 2006 Aug;173(4):2403-6. doi: 10.1534/genetics.106.059253. Epub 2006 May 15. Genetics. 2006. PMID: 16702418 Free PMC article. Review.
Cited by
-
Domain requirements of the JIL-1 tandem kinase for histone H3 serine 10 phosphorylation and chromatin remodeling in vivo.J Biol Chem. 2013 Jul 5;288(27):19441-9. doi: 10.1074/jbc.M113.464271. Epub 2013 May 30. J Biol Chem. 2013. PMID: 23723094 Free PMC article.
-
H3S10 phosphorylation by the JIL-1 kinase regulates H3K9 dimethylation and gene expression at the white locus in Drosophila.Fly (Austin). 2012 Apr-Jun;6(2):93-7. doi: 10.4161/fly.20029. Epub 2012 Apr 1. Fly (Austin). 2012. PMID: 22634714 Free PMC article.
-
The JIL-1 kinase affects telomere expression in the different telomere domains of Drosophila.PLoS One. 2013 Nov 14;8(11):e81543. doi: 10.1371/journal.pone.0081543. eCollection 2013. PLoS One. 2013. PMID: 24244743 Free PMC article.
-
Tethering of CHROMATOR and dCTCF proteins results in decompaction of condensed bands in the Drosophila melanogaster polytene chromosomes but does not affect their transcription and replication timing.PLoS One. 2018 Apr 2;13(4):e0192634. doi: 10.1371/journal.pone.0192634. eCollection 2018. PLoS One. 2018. PMID: 29608600 Free PMC article.
-
The effect of JIL-1 on position-effect variegation is proportional to the total amount of heterochromatin in the genome.Fly (Austin). 2013 Apr-Jun;7(2):129-33. doi: 10.4161/fly.24266. Epub 2013 Apr 1. Fly (Austin). 2013. PMID: 23579201 Free PMC article.
References
-
- Allis C. D., Jenuwein T., Reinberg D. (2007). Epigenetics. Cold Spring HarborNew York: Cold Sping Harbor Laboratory Press;
-
- Berger S. L. (2007). The complex language of chromatin regulation during transcription. Nature 447, 407-412 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
