

Characterization of Escherichia coli dinJ-yafQ toxin-antitoxin system using insights from mutagenesis data
- PMID: 22247505
- PMCID: PMC3294823
- DOI: 10.1128/JB.06104-11
Characterization of Escherichia coli dinJ-yafQ toxin-antitoxin system using insights from mutagenesis data
Abstract
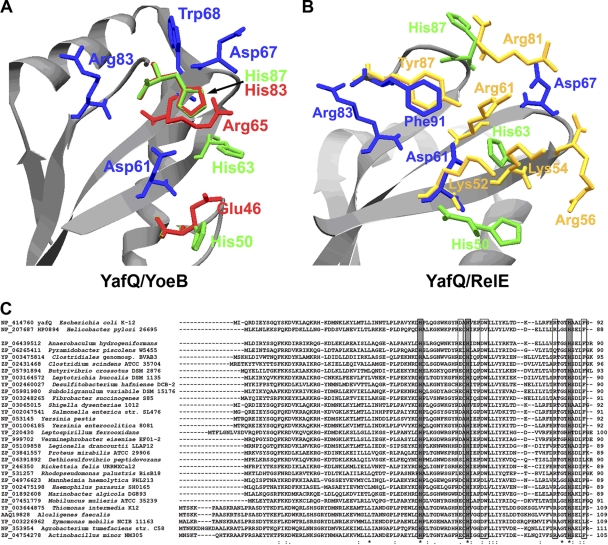
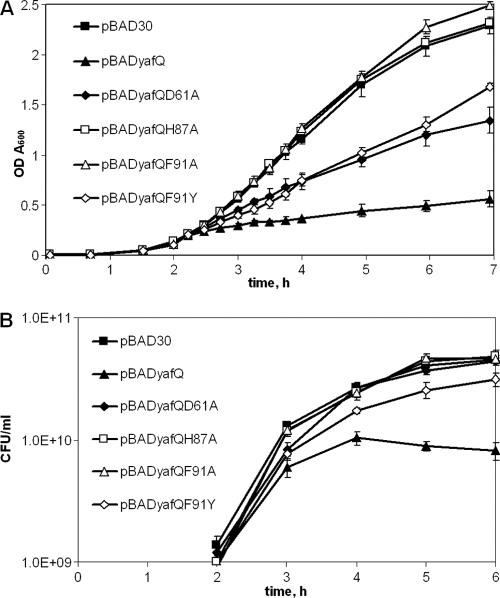
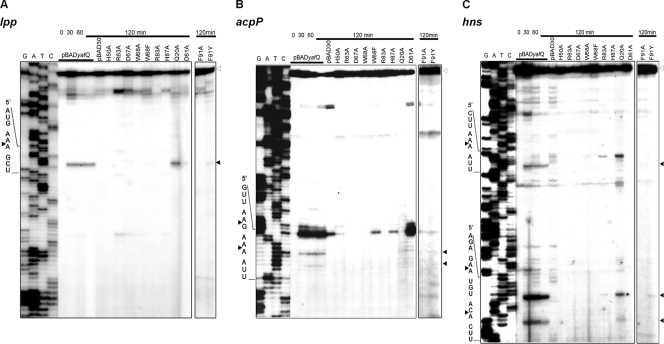
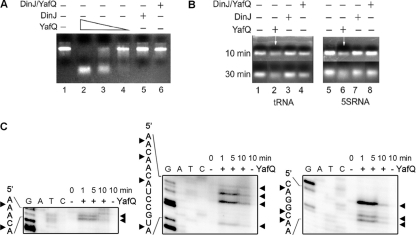
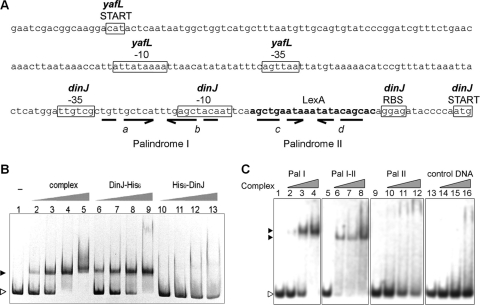
Escherichia coli dinJ-yafQ operon codes for a functional toxin-antitoxin (TA) system. YafQ toxin is an RNase which, upon overproduction, specifically inhibits the translation process by cleaving cellular mRNA at specific sequences. DinJ is an antitoxin and counteracts YafQ-mediated toxicity by forming a strong protein complex. In the present study we used site-directed mutagenesis of YafQ to determine the amino acids important for its catalytic activity. His50Ala, His63Ala, Asp67Ala, Trp68Ala, Trp68Phe, Arg83Ala, His87Ala, and Phe91Ala substitutions of the predicted active-site residues of YafQ abolished mRNA cleavage in vivo, whereas Asp61Ala and Phe91Tyr mutations inhibited YafQ RNase activity only moderately. We show that YafQ, upon overexpression, cleaved mRNAs preferably 5' to A between the second and third nucleotides in the codon in vivo. YafQ also showed RNase activity against mRNA, tRNA, and 5S rRNA molecules in vitro, albeit with no strong specificity. The endoribonuclease activity of YafQ was inhibited in the complex with DinJ antitoxin in vitro. DinJ-YafQ protein complex and DinJ antitoxin alone selectively bind to one of the two palindromic sequences present in the intergenic region upstream of the dinJ-yafQ operon, suggesting the autoregulation mode of this TA system.
Figures
References
-
- Arnold K, Bordoli L, Kopp J, Schwede T. 2006. The Swiss-Model Workspace: a web-based environment for protein structure homology modeling. Bioinformatics 22:195–201 - PubMed
-
- Bauerová-Hlinková V, Dvorský R, Perecko D, Povazanec F, Sevcík J. 2009. Structure of RNase Sa2 complexes with mononucleotides: new aspects of catalytic reaction and substrate recognition. FEBS J. 276:4156–4168 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
