

Merlin-deficient human tumors show loss of contact inhibition and activation of Wnt/β-catenin signaling linked to the PDGFR/Src and Rac/PAK pathways
- PMID: 22247700
- PMCID: PMC3257450
- DOI: 10.1593/neo.111060
Merlin-deficient human tumors show loss of contact inhibition and activation of Wnt/β-catenin signaling linked to the PDGFR/Src and Rac/PAK pathways
Abstract
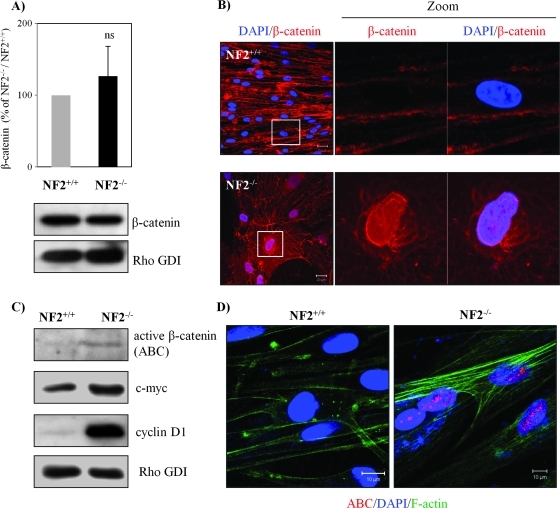
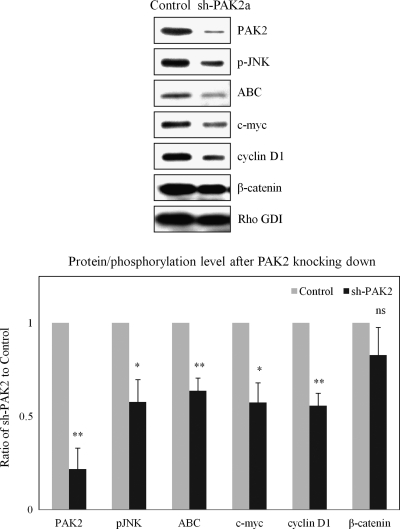
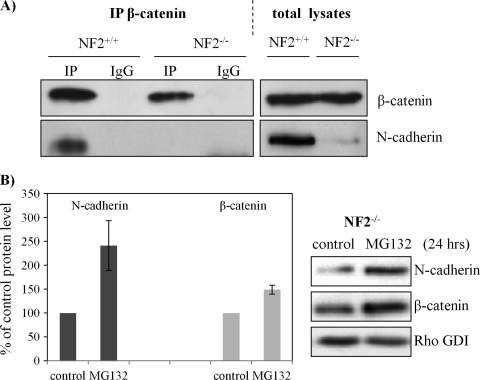
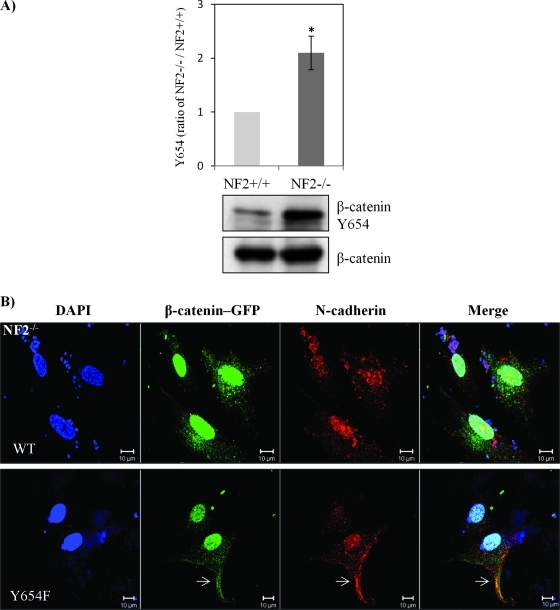
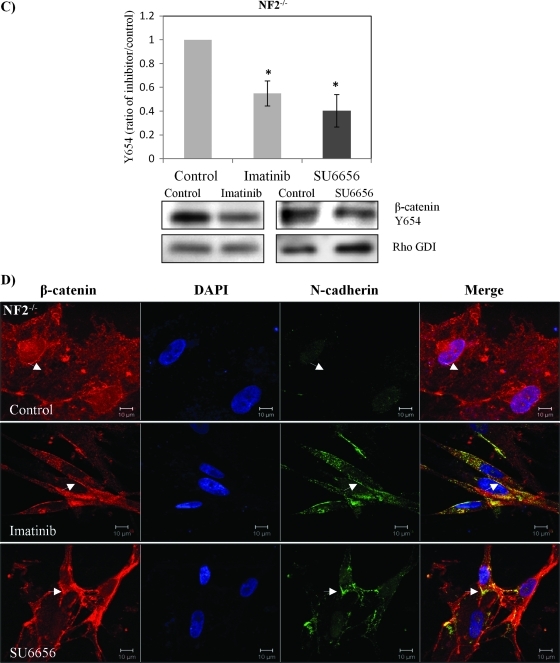
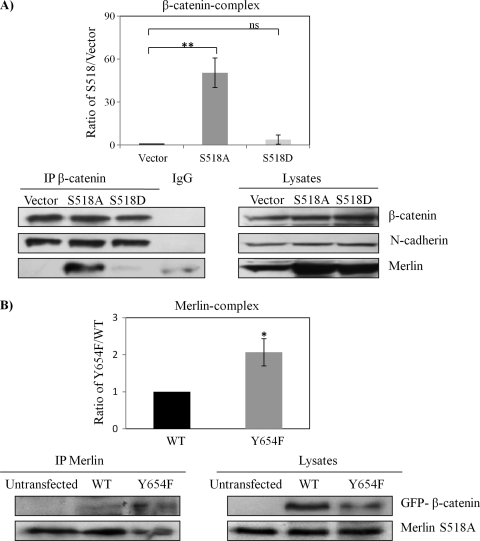
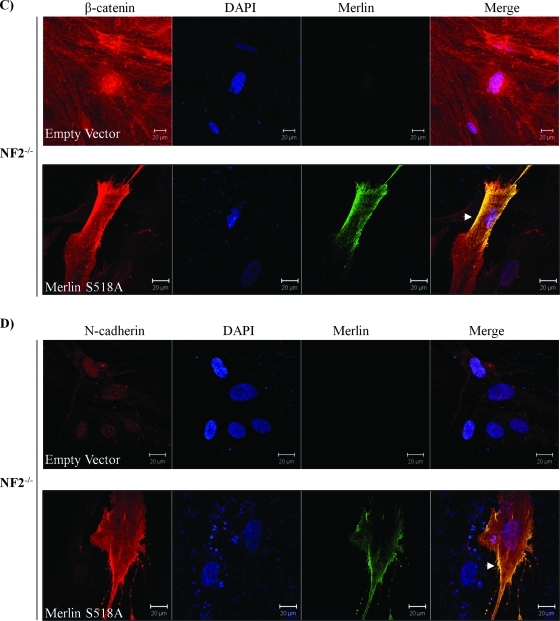
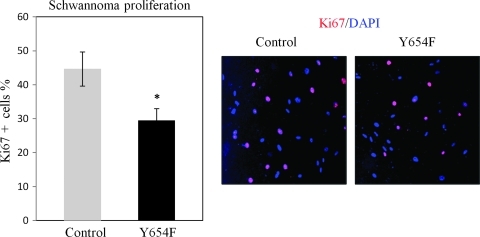
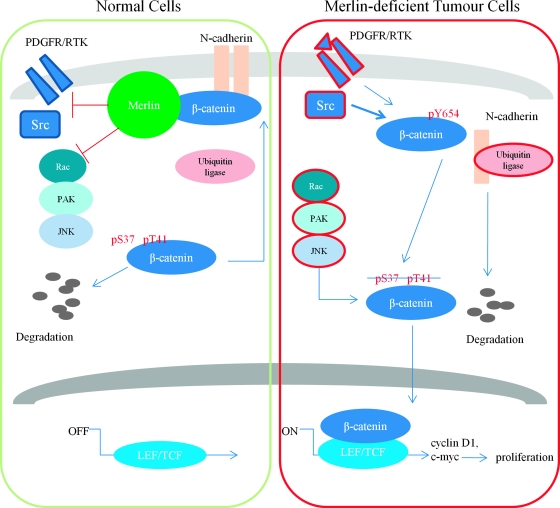
Neurofibromatosis type 2 (NF2) is an inherited predisposition cancer syndrome characterized by the development of multiple benign tumors in the nervous system including schwannomas, meningiomas, and ependymomas. Using a disease model comprising primary human schwannoma cells, we previously demonstrated that adherens junctions (AJs) are impaired in schwannoma cells because of a ubiquitous, upregulated Rac activity. However, the mechanism by which loss of contact inhibition leads to proliferation remains obscure in merlin-deficient tumors. In this study, we show that proliferative Wnt/β-catenin signaling is elevated as active β-catenin (dephosphorylated at serine 37 and threoine 41) localizes to the nucleus and the Wnt targets genes c-myc and cyclin D1 are upregulated in confluent human schwannoma cells. We demonstrate that Rac effector p21-activated kinase 2 (PAK2) is essential for the activation of Wnt/β-catenin signaling because depletion of PAK2 suppressed active β-catenin, c-myc, and cyclin D1. Most importantly, the link between the loss of the AJ complex and the increased proliferation in human schwannoma cells is connected by Src and platelet-derived growth factor receptor-induced tyrosine 654 phosphorylation on β-catenin and associated with degradation of N-cadherin. We also demonstrate that active merlin maintains β-catenin and N-cadherin complex at the plasma membrane through direct regulation. Finally, we demonstrate that phosphorylation of tyrosine 654 is critical for the increased proliferation in human schwannoma cells because overexpression of a Y654F mutant β-catenin reduces hyperproliferation of schwannoma cells. We suggest a model that these pathways are coordinated and relevant for proliferation in merlin-deficient tumors.
Figures
References
-
- Hanemann CO. Magic but treatable? Tumours due to loss of merlin. Brain. 2008;131:606–615. - PubMed
-
- Ammoun S, Flaiz C, Ristic N, Schuldt J, Hanemann CO. Dissecting and targeting the growth factor-dependent and growth factor-independent extracellular signal-regulated kinase pathway in human schwannoma. Cancer Res. 2008;68:5236–5245. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous