

CNGA3 is expressed in inner ear hair cells and binds to an intracellular C-terminus domain of EMILIN1
- PMID: 22248097
- PMCID: PMC6363478
- DOI: 10.1042/BJ20111255
CNGA3 is expressed in inner ear hair cells and binds to an intracellular C-terminus domain of EMILIN1
Abstract
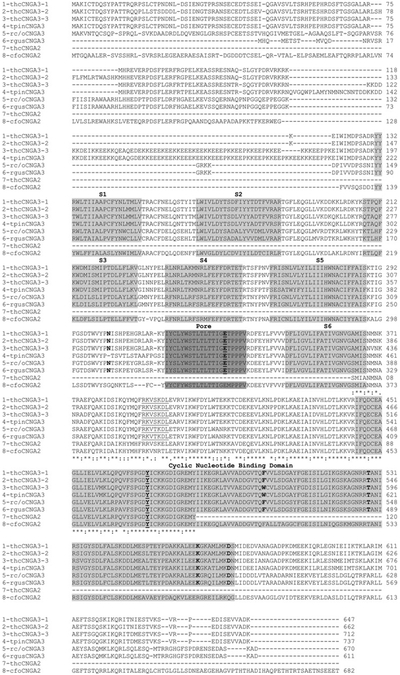
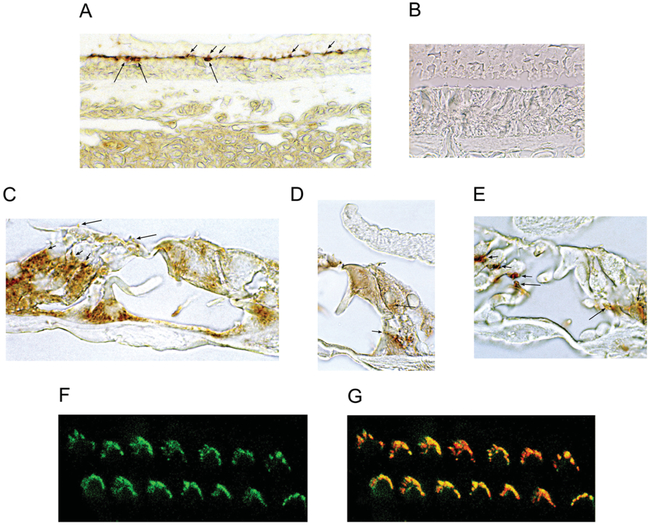
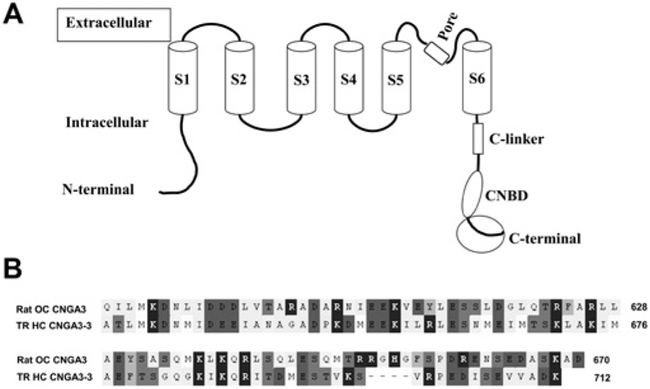
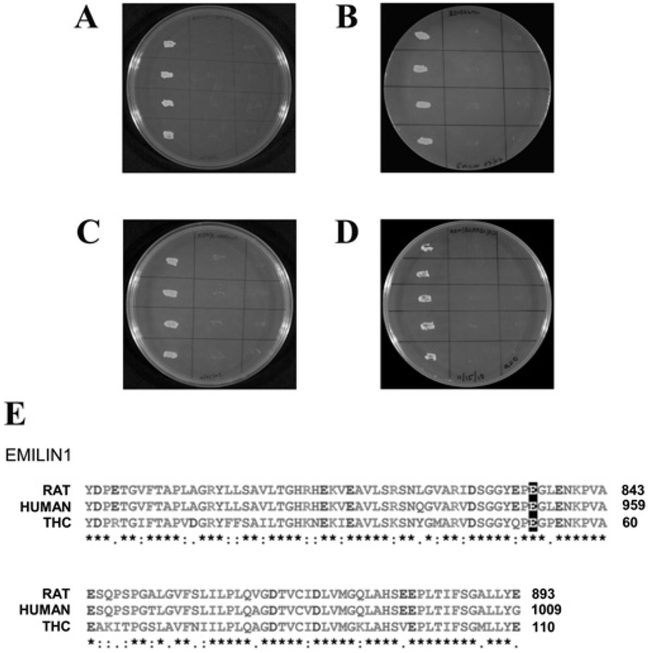
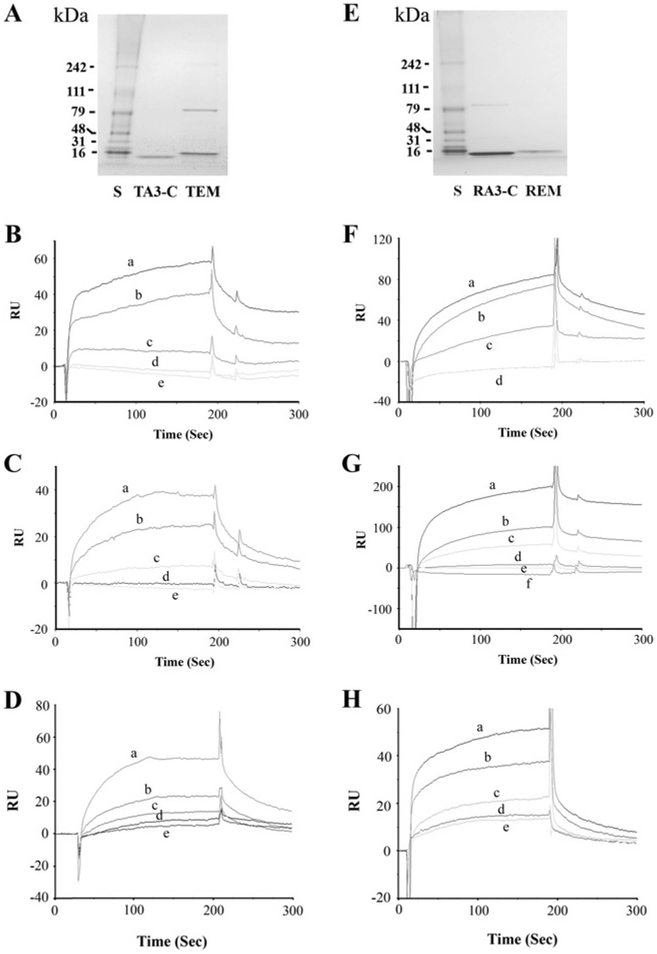
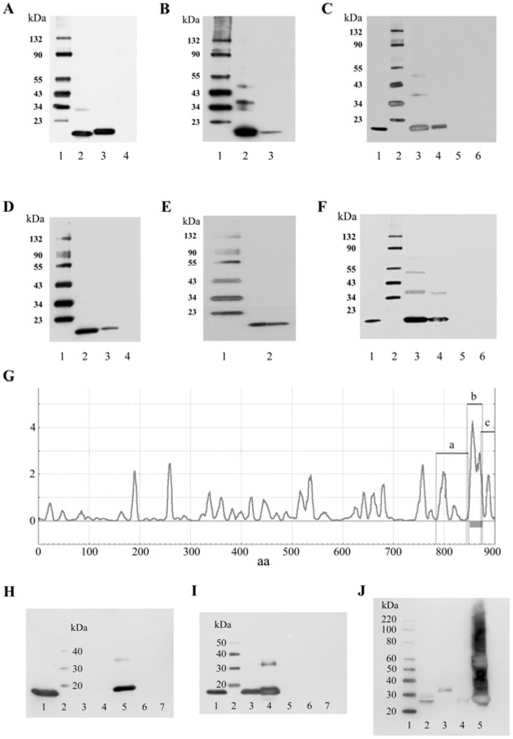
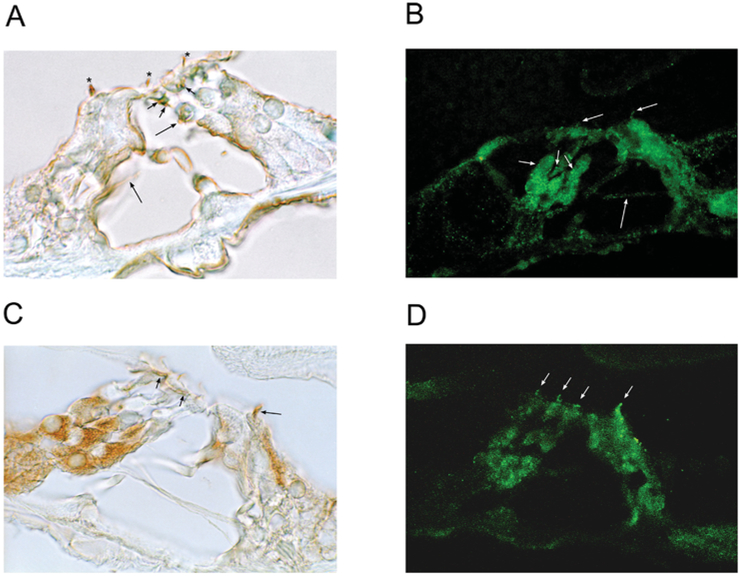
The molecular characteristics of CNG (cyclic nucleotide-gated) channels in auditory/vestibular hair cells are largely unknown, unlike those of CNG mediating sensory transduction in vision and olfaction. In the present study we report the full-length sequence for three CNGA3 variants in a hair cell preparation from the trout saccule with high identity to CNGA3 in olfactory receptor neurons/cone photoreceptors. A custom antibody targeting the N-terminal sequence immunolocalized CNGA3 to the stereocilia and subcuticular plate region of saccular hair cells. The cytoplasmic C-terminus of CNGA3 was found by yeast two-hybrid analysis to bind the C-terminus of EMILIN1 (elastin microfibril interface-located protein 1) in both the vestibular hair cell model and rat organ of Corti. Specific binding between CNGA3 and EMILIN1 was confirmed with surface plasmon resonance analysis, predicting dependence on Ca2+ with Kd=1.6×10-6 M for trout hair cell proteins and Kd=2.7×10-7 M for organ of Corti proteins at 68 μM Ca2+. Pull-down assays indicated that the binding to organ of Corti CNGA3 was attributable to the EMILIN1 intracellular sequence that follows a predicted transmembrane domain in the C-terminus. Saccular hair cells also express the transcript for PDE6C (phosphodiesterase 6C), which in cone photoreceptors regulates the degradation of cGMP used to gate CNGA3 in phototransduction. Taken together, the evidence supports the existence in saccular hair cells of a molecular pathway linking CNGA3, its binding partner EMILIN1 (and β1 integrin) and cGMP-specific PDE6C, which is potentially replicated in cochlear outer hair cells, given stereociliary immunolocalizations of CNGA3, EMILIN1 and PDE6C.
Figures

Similar articles
-
HCN1 and HCN2 proteins are expressed in cochlear hair cells: HCN1 can form a ternary complex with protocadherin 15 CD3 and F-actin-binding filamin A or can interact with HCN2.J Biol Chem. 2012 Nov 2;287(45):37628-46. doi: 10.1074/jbc.M112.375832. Epub 2012 Sep 4. J Biol Chem. 2012. PMID: 22948144 Free PMC article.
-
Cyclic nucleotide-gated channel α-3 (CNGA3) interacts with stereocilia tip-link cadherin 23 + exon 68 or alternatively with myosin VIIa, two proteins required for hair cell mechanotransduction.J Biol Chem. 2013 Mar 8;288(10):7215-29. doi: 10.1074/jbc.M112.443226. Epub 2013 Jan 17. J Biol Chem. 2013. PMID: 23329832 Free PMC article.
-
Calcium-dependent binding of HCN1 channel protein to hair cell stereociliary tip link protein protocadherin 15 CD3.J Biol Chem. 2009 Jan 30;284(5):3227-3238. doi: 10.1074/jbc.M806177200. Epub 2008 Nov 13. J Biol Chem. 2009. PMID: 19008224 Free PMC article.
-
Retinal Cyclic Nucleotide-Gated Channels: From Pathophysiology to Therapy.Int J Mol Sci. 2018 Mar 7;19(3):749. doi: 10.3390/ijms19030749. Int J Mol Sci. 2018. PMID: 29518895 Free PMC article. Review.
-
Cyclic nucleotide-gated channels.Handb Exp Pharmacol. 2009;(191):111-36. doi: 10.1007/978-3-540-68964-5_7. Handb Exp Pharmacol. 2009. PMID: 19089328 Review.
Cited by
-
HCN1 and HCN2 proteins are expressed in cochlear hair cells: HCN1 can form a ternary complex with protocadherin 15 CD3 and F-actin-binding filamin A or can interact with HCN2.J Biol Chem. 2012 Nov 2;287(45):37628-46. doi: 10.1074/jbc.M112.375832. Epub 2012 Sep 4. J Biol Chem. 2012. PMID: 22948144 Free PMC article.
-
Cyclic nucleotide-gated channel α-3 (CNGA3) interacts with stereocilia tip-link cadherin 23 + exon 68 or alternatively with myosin VIIa, two proteins required for hair cell mechanotransduction.J Biol Chem. 2013 Mar 8;288(10):7215-29. doi: 10.1074/jbc.M112.443226. Epub 2013 Jan 17. J Biol Chem. 2013. PMID: 23329832 Free PMC article.
-
Dopamine D1A directly interacts with otoferlin synaptic pathway proteins: Ca2+ and phosphorylation underlie an NSF-to-AP2mu1 molecular switch.Biochem J. 2017 Jan 1;474(1):79-104. doi: 10.1042/BCJ20160690. Epub 2016 Nov 7. Biochem J. 2017. PMID: 27821621 Free PMC article.
-
Diabetic Aldehyde Dehydrogenase 2 Mutant (ALDH2*2) Mice Are More Susceptible to Cardiac Ischemic-Reperfusion Injury Due to 4-Hydroxy-2-Nonenal Induced Coronary Endothelial Cell Damage.J Am Heart Assoc. 2021 Sep 21;10(18):e021140. doi: 10.1161/JAHA.121.021140. Epub 2021 Sep 6. J Am Heart Assoc. 2021. PMID: 34482710 Free PMC article.
-
QTL Mapping of Endocochlear Potential Differences between C57BL/6J and BALB/cJ mice.J Assoc Res Otolaryngol. 2016 Jun;17(3):173-94. doi: 10.1007/s10162-016-0558-8. Epub 2016 Mar 15. J Assoc Res Otolaryngol. 2016. PMID: 26980469 Free PMC article.
References
-
- Zagotta WN and Siegelbaum SA (1996) Structure and function of cyclic nucleotide-gated channels. Annu. Rev. Neurosci 19, 235–263 - PubMed
-
- Biel M and Michalakis S (2009) Cyclic nucleotide-gated channels. Handb. Exp. Pharmacol 191, 111–136 - PubMed
-
- Misaka T, Kusakabe Y, Emori Y, Gonoi T, Arai S and Abe K (1997) Taste buds have a cyclic nucleotide-activated channel, CNGgust. J. Biol. Chem 272, 22623–22629 - PubMed
-
- Lee HM, Park YS, Kim W and Park CS (2001) Electrophysiological characteristics of rat gustatory cyclic nucleotide-gated channel expressed in Xenopus oocytes. J. Neurophysiol 85, 2335–2349 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous