

Defects in leaf carbohydrate metabolism compromise acclimation to high light and lead to a high chlorophyll fluorescence phenotype in Arabidopsis thaliana
- PMID: 22248311
- PMCID: PMC3353854
- DOI: 10.1186/1471-2229-12-8
Defects in leaf carbohydrate metabolism compromise acclimation to high light and lead to a high chlorophyll fluorescence phenotype in Arabidopsis thaliana
Abstract
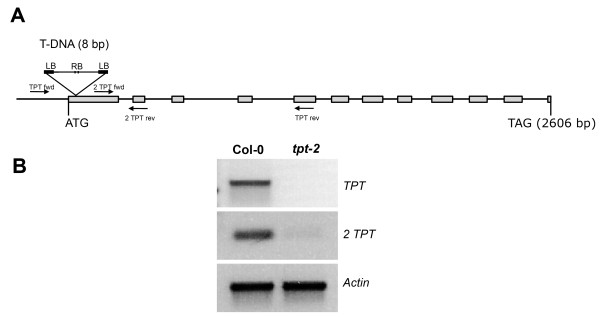
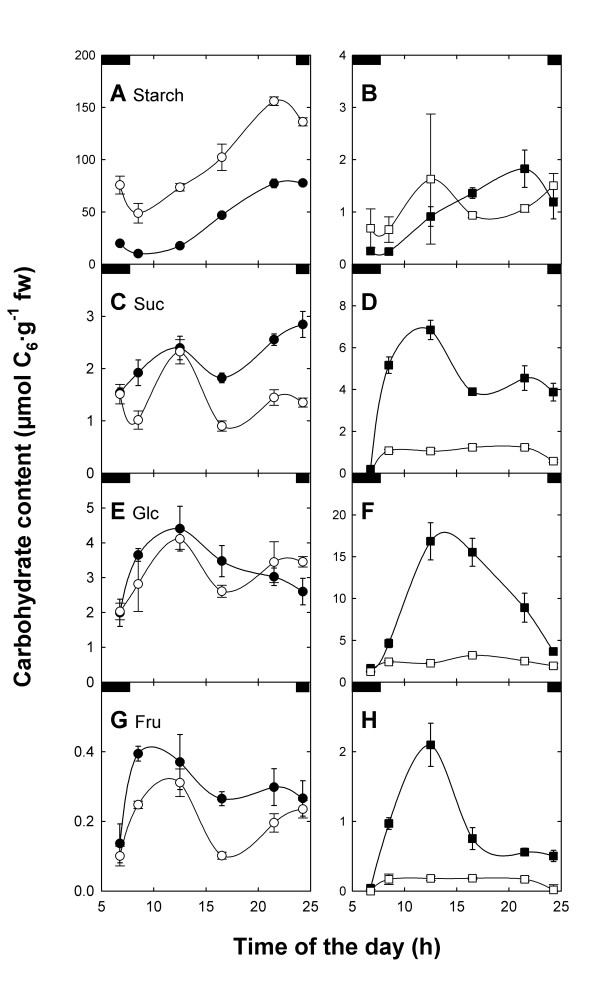
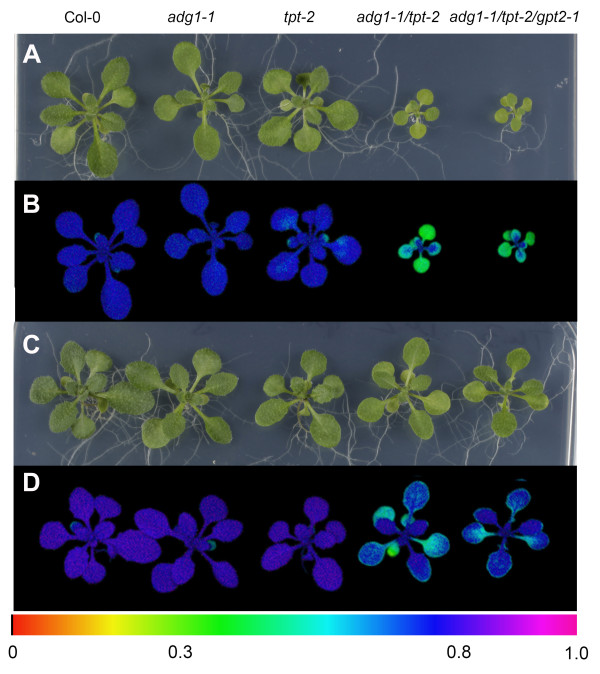
Background: We have studied the impact of carbohydrate-starvation on the acclimation response to high light using Arabidopsis thaliana double mutants strongly impaired in the day- and night path of photoassimilate export from the chloroplast. A complete knock-out mutant of the triose phosphate/phosphate translocator (TPT; tpt-2 mutant) was crossed to mutants defective in (i) starch biosynthesis (adg1-1, pgm1 and pgi1-1; knock-outs of ADP-glucose pyrophosphorylase, plastidial phosphoglucomutase and phosphoglucose isomerase) or (ii) starch mobilization (sex1-3, knock-out of glucan water dikinase) as well as in (iii) maltose export from the chloroplast (mex1-2).
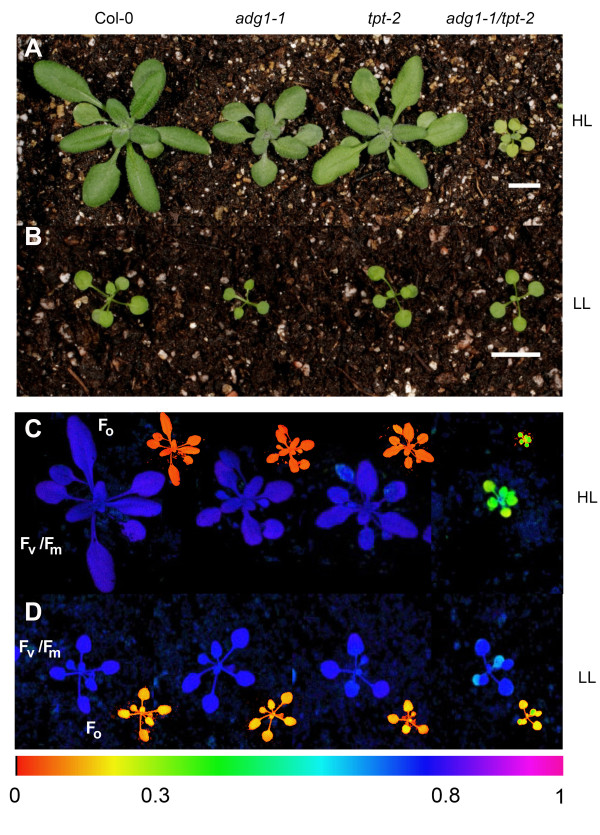
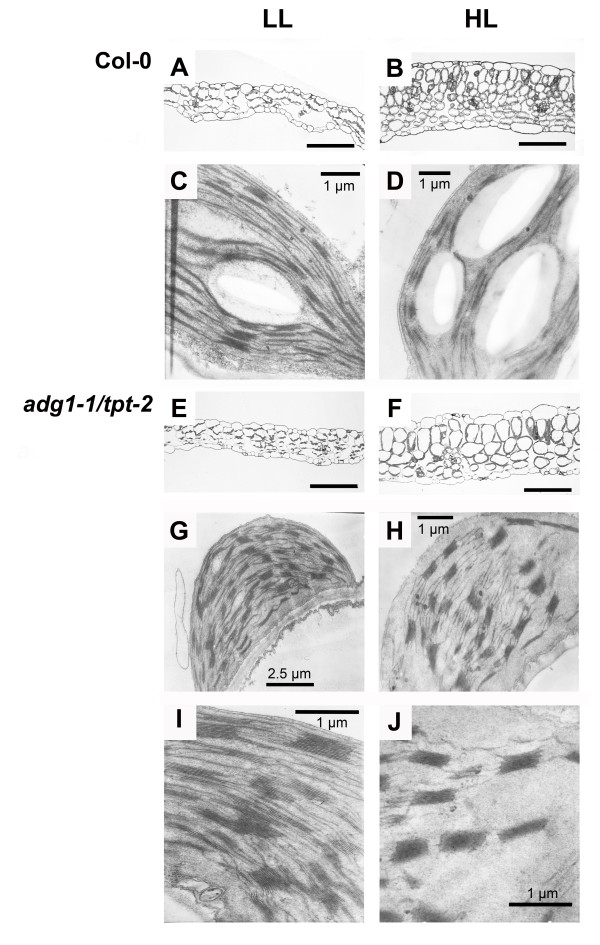
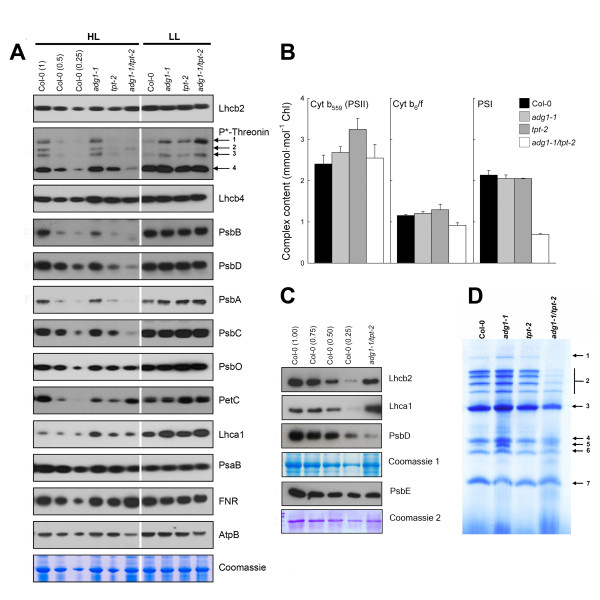
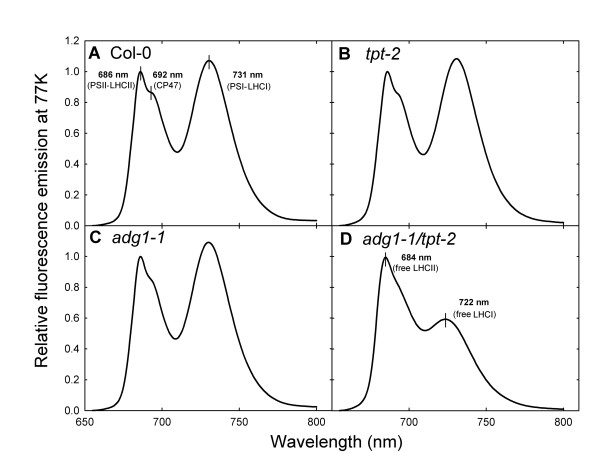
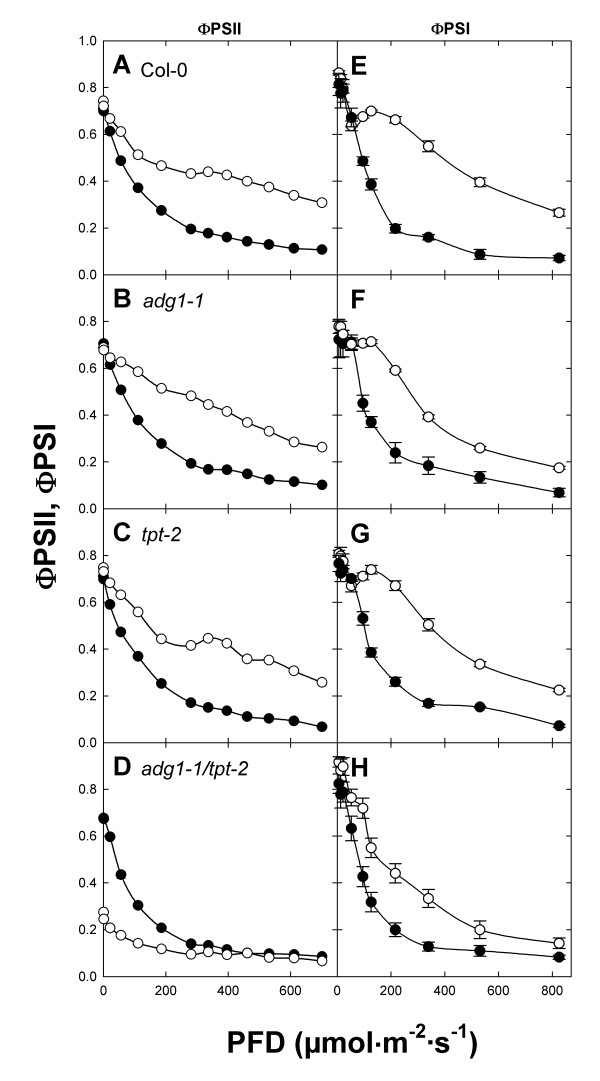
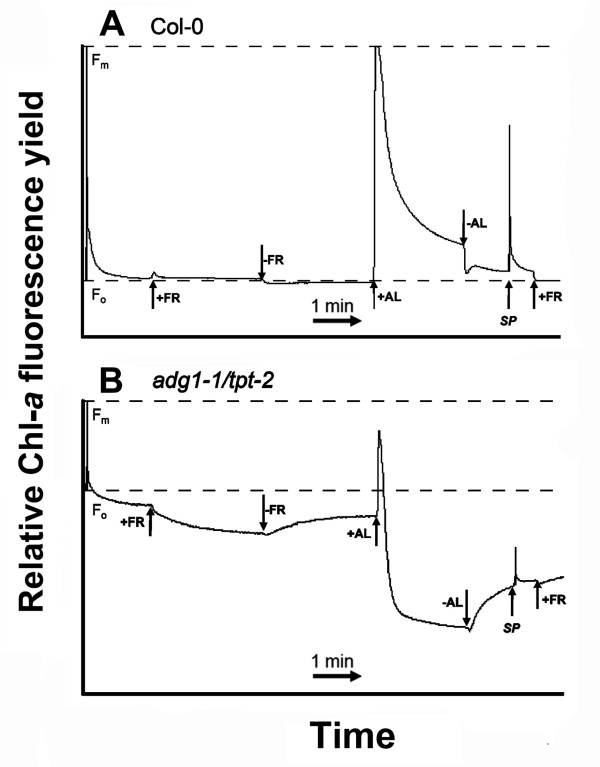
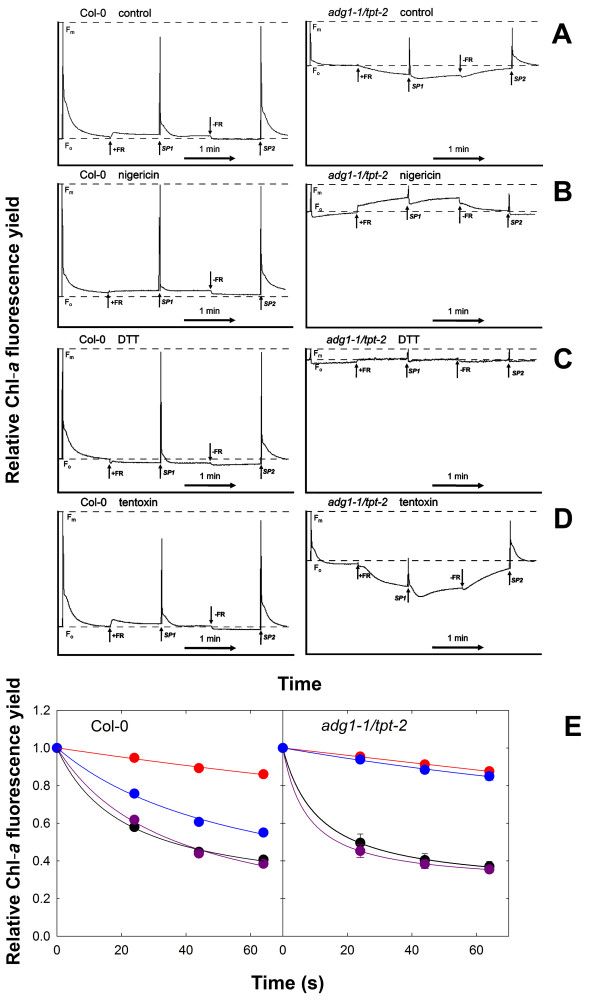
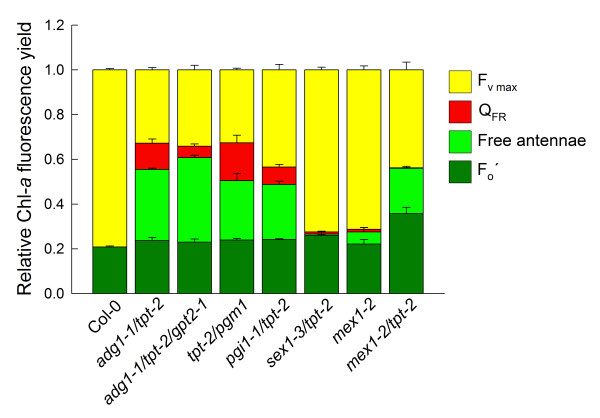
Results: All double mutants were viable and indistinguishable from the wild type when grown under low light conditions, but--except for sex1-3/tpt-2--developed a high chlorophyll fluorescence (HCF) phenotype and growth retardation when grown in high light. Immunoblots of thylakoid proteins, Blue-Native gel electrophoresis and chlorophyll fluorescence emission analyses at 77 Kelvin with the adg1-1/tpt-2 double mutant revealed that HCF was linked to a specific decrease in plastome-encoded core proteins of both photosystems (with the exception of the PSII component cytochrome b559), whereas nuclear-encoded antennae (LHCs) accumulated normally, but were predominantly not attached to their photosystems. Uncoupled antennae are the major cause for HCF of dark-adapted plants. Feeding of sucrose or glucose to high light-grown adg1-1/tpt-2 plants rescued the HCF- and growth phenotypes. Elevated sugar levels induce the expression of the glucose-6-phosphate/phosphate translocator2 (GPT2), which in principle could compensate for the deficiency in the TPT. A triple mutant with an additional defect in GPT2 (adg1-1/tpt-2/gpt2-1) exhibited an identical rescue of the HCF- and growth phenotype in response to sugar feeding as the adg1-1/tpt-2 double mutant, indicating that this rescue is independent from the sugar-triggered induction of GPT2.
Conclusions: We propose that cytosolic carbohydrate availability modulates acclimation to high light in A. thaliana. It is conceivable that the strong relationship between the chloroplast and nucleus with respect to a co-ordinated expression of photosynthesis genes is modified in carbohydrate-starved plants. Hence carbohydrates may be considered as a novel component involved in chloroplast-to-nucleus retrograde signaling, an aspect that will be addressed in future studies.
Figures
Similar articles
-
The essential role of sugar metabolism in the acclimation response of Arabidopsis thaliana to high light intensities.J Exp Bot. 2014 Apr;65(6):1619-36. doi: 10.1093/jxb/eru027. Epub 2014 Feb 12. J Exp Bot. 2014. PMID: 24523502 Free PMC article.
-
The Mysterious Rescue of adg1-1/tpt-2 - an Arabidopsis thaliana Double Mutant Impaired in Acclimation to High Light - by Exogenously Supplied Sugars.Front Plant Sci. 2012 Nov 30;3:265. doi: 10.3389/fpls.2012.00265. eCollection 2012. Front Plant Sci. 2012. PMID: 23233856 Free PMC article.
-
The role of plastidial glucose-6-phosphate/phosphate translocators in vegetative tissues of Arabidopsis thaliana mutants impaired in starch biosynthesis.Plant Biol (Stuttg). 2010 Sep;12 Suppl 1:115-28. doi: 10.1111/j.1438-8677.2010.00349.x. Plant Biol (Stuttg). 2010. PMID: 20712627
-
How sugars might coordinate chloroplast and nuclear gene expression during acclimation to high light intensities.Mol Plant. 2014 Jul;7(7):1121-37. doi: 10.1093/mp/ssu064. Epub 2014 May 23. Mol Plant. 2014. PMID: 25006007 Review.
-
The functions of WHIRLY1 and REDOX-RESPONSIVE TRANSCRIPTION FACTOR 1 in cross tolerance responses in plants: a hypothesis.Philos Trans R Soc Lond B Biol Sci. 2014 Mar 3;369(1640):20130226. doi: 10.1098/rstb.2013.0226. Print 2014 Apr 19. Philos Trans R Soc Lond B Biol Sci. 2014. PMID: 24591713 Free PMC article. Review.
Cited by
-
Dynamic Acclimation to High Light in Arabidopsis thaliana Involves Widespread Reengineering of the Leaf Proteome.Front Plant Sci. 2017 Jul 20;8:1239. doi: 10.3389/fpls.2017.01239. eCollection 2017. Front Plant Sci. 2017. PMID: 28775726 Free PMC article.
-
Reduced vacuolar β-1,3-glucan synthesis affects carbohydrate metabolism as well as plastid homeostasis and structure in Phaeodactylum tricornutum.Proc Natl Acad Sci U S A. 2018 May 1;115(18):4791-4796. doi: 10.1073/pnas.1719274115. Epub 2018 Apr 18. Proc Natl Acad Sci U S A. 2018. PMID: 29669920 Free PMC article.
-
The essential role of sugar metabolism in the acclimation response of Arabidopsis thaliana to high light intensities.J Exp Bot. 2014 Apr;65(6):1619-36. doi: 10.1093/jxb/eru027. Epub 2014 Feb 12. J Exp Bot. 2014. PMID: 24523502 Free PMC article.
-
An Arabidopsis mutant with high operating efficiency of Photosystem II and low chlorophyll fluorescence.Sci Rep. 2017 Jun 12;7(1):3314. doi: 10.1038/s41598-017-03611-1. Sci Rep. 2017. PMID: 28607440 Free PMC article.
-
The availability of neither D2 nor CP43 limits the biogenesis of photosystem II in tobacco.Plant Physiol. 2021 Apr 2;185(3):1111-1130. doi: 10.1093/plphys/kiaa052. Plant Physiol. 2021. PMID: 33793892 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
