

Ceramide synthases at the centre of sphingolipid metabolism and biology
- PMID: 22248339
- PMCID: PMC3689921
- DOI: 10.1042/BJ20111626
Ceramide synthases at the centre of sphingolipid metabolism and biology
Abstract
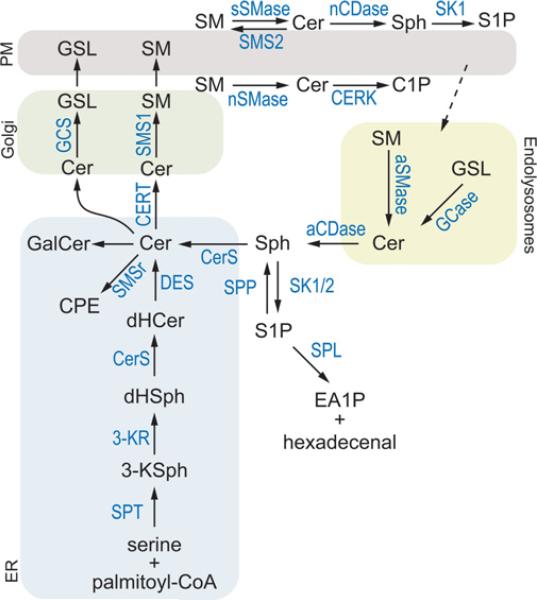
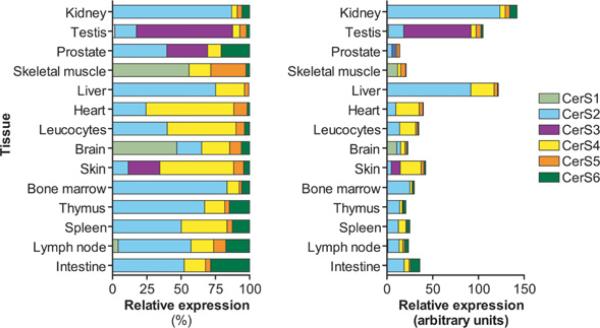
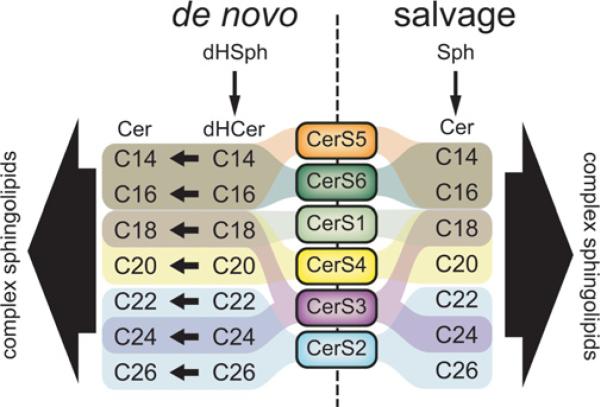
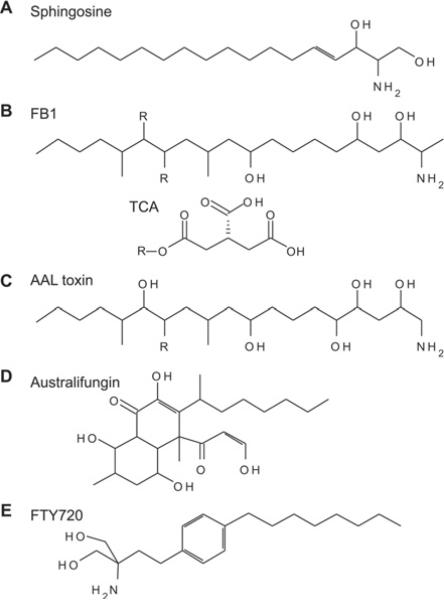
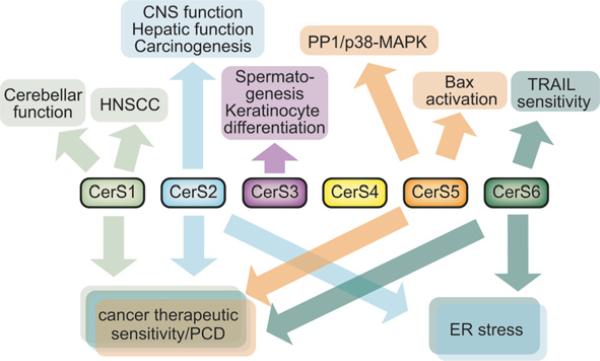
Sphingolipid metabolism in metazoan cells consists of a complex interconnected web of numerous enzymes, metabolites and modes of regulation. At the centre of sphingolipid metabolism reside CerSs (ceramide synthases), a group of enzymes that catalyse the formation of ceramides from sphingoid base and acyl-CoA substrates. From a metabolic perspective, these enzymes occupy a unique niche in that they simultaneously regulate de novo sphingolipid synthesis and the recycling of free sphingosine produced from the degradation of pre-formed sphingolipids (salvage pathway). Six mammalian CerSs (CerS1-CerS6) have been identified. Unique characteristics have been described for each of these enzymes, but perhaps the most notable is the ability of individual CerS isoforms to produce ceramides with characteristic acyl-chain distributions. Through this control of acyl-chain length and perhaps in a compartment-specific manner, CerSs appear to regulate multiple aspects of sphingolipid-mediated cell and organismal biology. In the present review, we discuss the function of CerSs as critical regulators of sphingolipid metabolism, highlight their unique characteristics and explore the emerging roles of CerSs in regulating programmed cell death, cancer and many other aspects of biology.
Figures
References
-
- Hannun YA, Obeid LM. Principles of bioactive lipid signalling: lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008;9:139–150. - PubMed
-
- Gatt S. Enzymic hydrolysis and synthesis of ceramides. J. Biol. Chem. 1963;238:3131–3133. - PubMed
-
- Gatt S. Enzymatic hydrolysis of sphingolipids. I. Hydrolysis and synthesis of ceramides by an enzyme from rat brain. J. Biol. Chem. 1966;241:3724–3730. - PubMed
-
- Yavin E, Gatt S. Enzymatic hydrolysis of sphingolipids. 8. Further purification and properties of rat brain ceramidase. Biochemistry. 1969;8:1692–1698. - PubMed
-
- Sribney M. Enzymatic synthesis of ceramide. Biochim. Biophys. Acta. 1966;125:542–547. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases