

Selenium is a modulator of circadian clock that protects mice from the toxicity of a chemotherapeutic drug via upregulation of the core clock protein, BMAL1
- PMID: 22249125
- PMCID: PMC3282084
- DOI: 10.18632/oncotarget.411
Selenium is a modulator of circadian clock that protects mice from the toxicity of a chemotherapeutic drug via upregulation of the core clock protein, BMAL1
Abstract
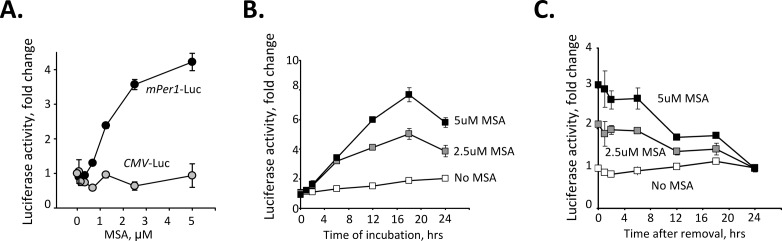
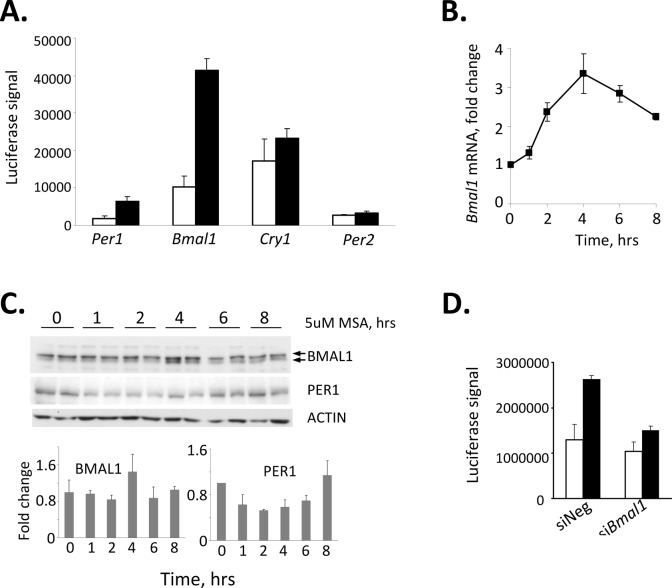
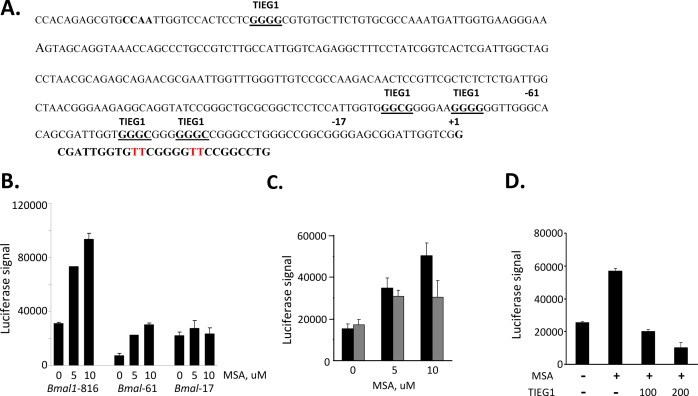
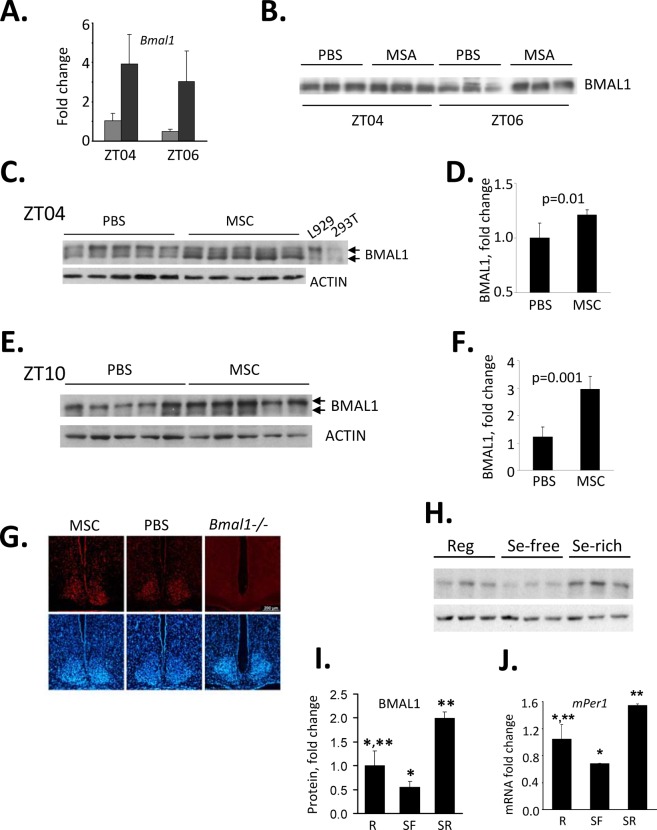
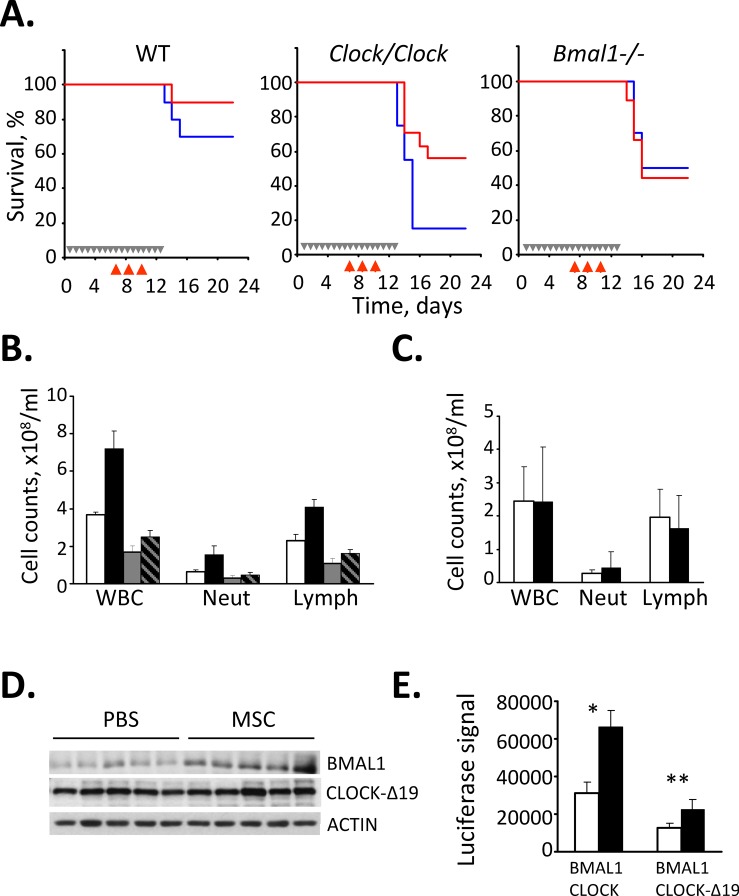
Selenium compounds are known as cancer preventive agents and are also able to ameliorate the toxicity associated with anti-cancer radiation and chemotherapy in mouse models. Sensitivity to the toxicity of chemotherapy is also modulated by the circadian clock, molecular time-keeping system that underlie daily fluctuations in multiple physiological and biochemical processes. Here we show that these two mechanisms are interconnected. By screening a library of small molecules in a cell-based reporter system, we identified L-methyl-selenocysteine as a positive regulator of the core clock protein, BMAL1. L-methyl-selenocysteine up-regulates BMAL1 at the transcriptional level both in cultured cells and in mice. We also show that in tissue culture selenium exerts its action by interfering with TIEG1-mediated repression of Bmal1 promoter. Selenium treatment fails to protect BMAL1-deficient mice from toxicity induced by the chemotherapeutic agent cyclophosphamide but does protect Clock mutant mice deficient in circadian rhythm control but having normal BMAL1. These findings define selenium as circadian modulator and indicate that the tissue protective effect of selenium results, at least in part, from up-regulation of BMAL1 expression and subsequent enhancement of CLOCK/BMAL1-mediated transcription.
Figures
Similar articles
-
PI3K regulates BMAL1/CLOCK-mediated circadian transcription from the Dbp promoter.Biosci Biotechnol Biochem. 2016 Jun;80(6):1131-40. doi: 10.1080/09168451.2015.1136885. Epub 2016 Mar 29. Biosci Biotechnol Biochem. 2016. PMID: 27022680
-
Disruption of CLOCK-BMAL1 transcriptional activity is responsible for aryl hydrocarbon receptor-mediated regulation of Period1 gene.Toxicol Sci. 2010 May;115(1):98-108. doi: 10.1093/toxsci/kfq022. Epub 2010 Jan 27. Toxicol Sci. 2010. PMID: 20106950 Free PMC article.
-
Transcriptional repressor TIEG1 regulates Bmal1 gene through GC box and controls circadian clockwork.Genes Cells. 2010 Feb;15(2):111-21. doi: 10.1111/j.1365-2443.2009.01371.x. Epub 2010 Jan 13. Genes Cells. 2010. PMID: 20070857
-
Circadian modification network of a core clock driver BMAL1 to harmonize physiology from brain to peripheral tissues.Neurochem Int. 2018 Oct;119:11-16. doi: 10.1016/j.neuint.2017.12.013. Epub 2018 Jan 3. Neurochem Int. 2018. PMID: 29305918 Review.
-
BMAL1 dephosphorylation determines the pace of the circadian clock.Genes Dev. 2021 Aug 1;35(15-16):1076-1078. doi: 10.1101/gad.348801.121. Genes Dev. 2021. PMID: 34341001 Free PMC article. Review.
Cited by
-
At the Interface of Lifestyle, Behavior, and Circadian Rhythms: Metabolic Implications.Front Nutr. 2019 Aug 28;6:132. doi: 10.3389/fnut.2019.00132. eCollection 2019. Front Nutr. 2019. PMID: 31555652 Free PMC article. Review.
-
Evolving roles of circadian rhythms in liver homeostasis and pathology.Oncotarget. 2016 Feb 23;7(8):8625-39. doi: 10.18632/oncotarget.7065. Oncotarget. 2016. PMID: 26843619 Free PMC article. Review.
-
Development and Therapeutic Potential of Small-Molecule Modulators of Circadian Systems.Annu Rev Pharmacol Toxicol. 2018 Jan 6;58:231-252. doi: 10.1146/annurev-pharmtox-010617-052645. Epub 2017 Oct 2. Annu Rev Pharmacol Toxicol. 2018. PMID: 28968186 Free PMC article. Review.
-
Multi-Modal Regulation of Circadian Physiology by Interactive Features of Biological Clocks.Biology (Basel). 2021 Dec 24;11(1):21. doi: 10.3390/biology11010021. Biology (Basel). 2021. PMID: 35053019 Free PMC article. Review.
-
Enhancement of NAD⁺-dependent SIRT1 deacetylase activity by methylselenocysteine resets the circadian clock in carcinogen-treated mammary epithelial cells.Oncotarget. 2015 Dec 15;6(40):42879-91. doi: 10.18632/oncotarget.6002. Oncotarget. 2015. PMID: 26544624 Free PMC article.
References
-
- Rayman MP. The importance of selenium to human health. Lancet. 2000;356:233–241. - PubMed
-
- Rayman MP. Selenium in cancer prevention: a review of the evidence and mechanism of action. Proc Nutr Soc. 2005;64:527–542. - PubMed
-
- Zeng H, Combs GF., Jr Selenium as an anticancer nutrient: roles in cell proliferation and tumor cell invasion. J Nutr Biochem. 2008;19:1–7. - PubMed
-
- Bhattacharya A, Toth K, Durrani FA, Cao S, Slocum HK, Chintala S, Rustum YM. Hypoxia-specific drug tirapazamine does not abrogate hypoxic tumor cells in combination therapy with irinotecan and methylselenocysteine in well-differentiated human head and neck squamous cell carcinoma a253 xenografts. Neoplasia. 2008;10:857–865. - PMC - PubMed
-
- Cao S, Durrani FA, Rustum YM. Selective modulation of the therapeutic efficacy of anticancer drugs by selenium containing compounds against human tumor xenografts. Clin Cancer Res. 2004;10:2561–2569. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
