

The B-cell superantigen Finegoldia magna protein L causes pulmonary inflammation by a mechanism dependent on MyD88 but not B cells or immunoglobulins
- PMID: 22249932
- PMCID: PMC3279619
- DOI: 10.1007/s00011-012-0436-8
The B-cell superantigen Finegoldia magna protein L causes pulmonary inflammation by a mechanism dependent on MyD88 but not B cells or immunoglobulins
Abstract
Objective and design: To determine whether Finegoldia magna protein L (PL) causes lung inflammation and, if so, whether the response is dependent on its immunoglobulin (Ig)-binding B-cell superantigenic property.
Material: Pulmonary inflammatory reactions were analyzed at various time points after intratracheal administration of PL to various strains of mice.
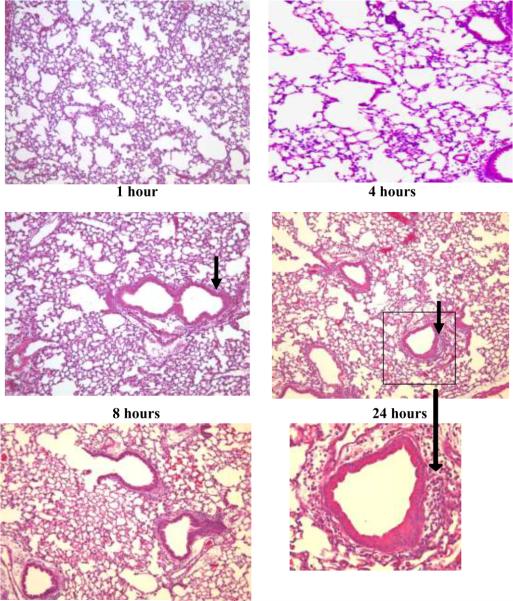
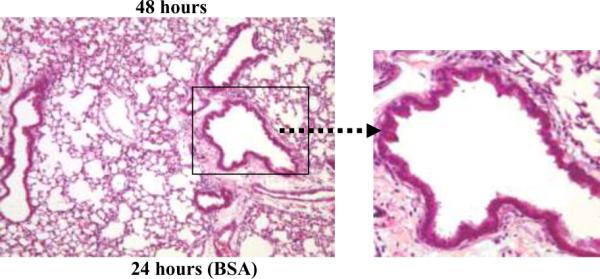
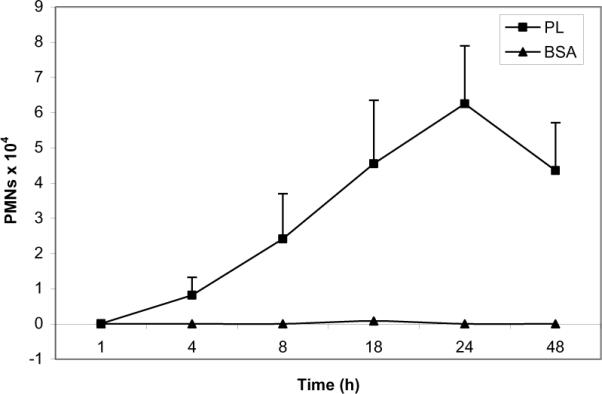
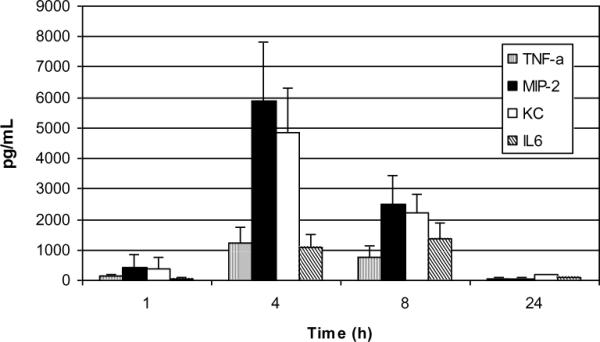
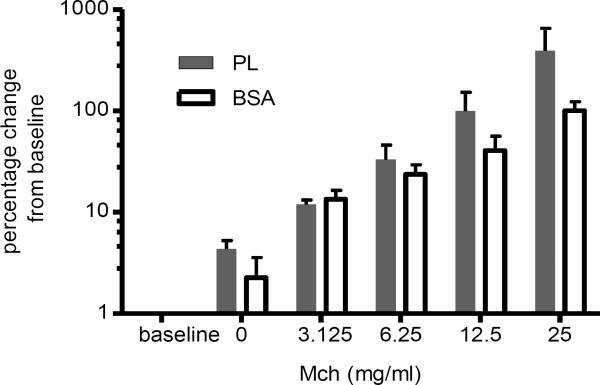
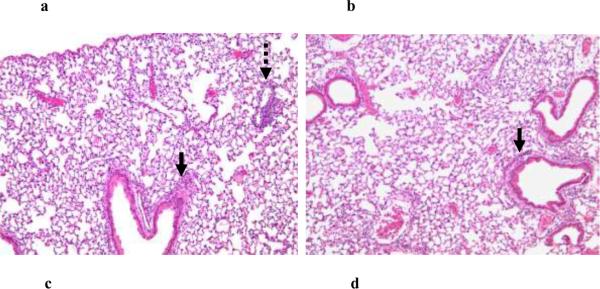
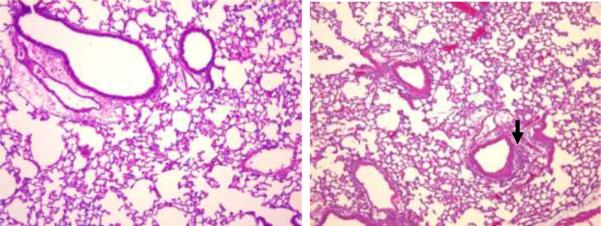
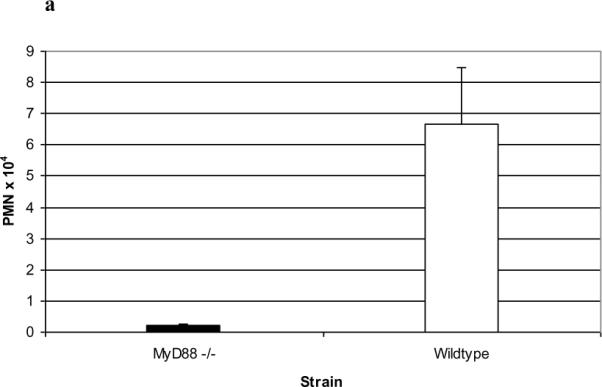
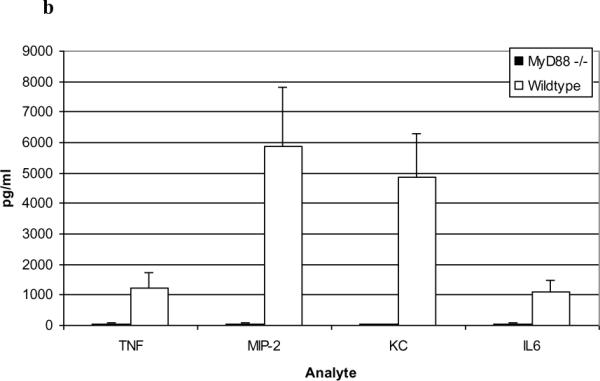
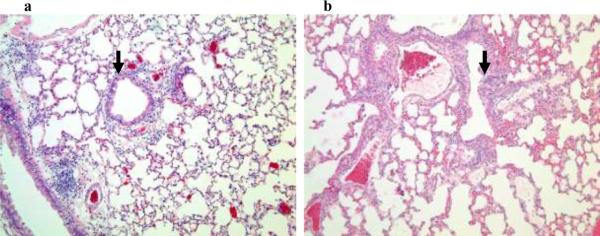
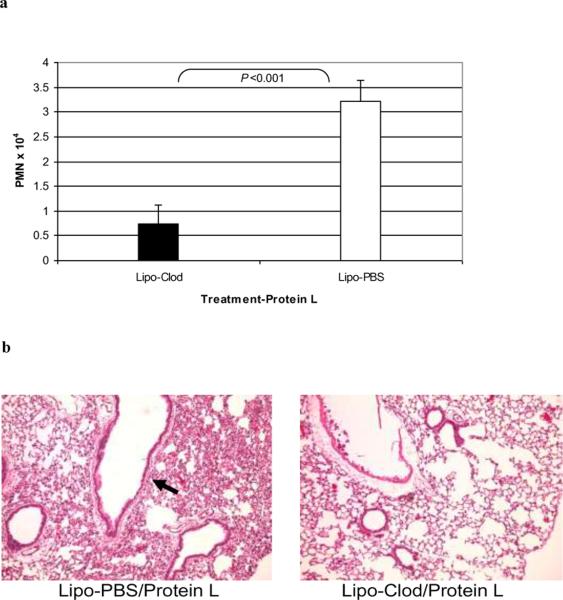
Results: PL caused peribronchial and perivascular inflammation that peaked at 18-24 h. Polymorphonuclear cells (PMNs) began to accumulate in bronchoalveolar lavage fluid (BALF) of PL-challenged mice by 4 h and accounted for >90% of leukocytes by 18-24 h. Inflammation was marked by the appearance of MIP-2, KC, TNF-α, and IL-6 in the BALF with peak levels attained 4 h after PL administration. PL-induced pulmonary inflammation was associated with increased airway hyper-reactivity following inhalation of methacholine. The inflammatory reaction was unabated in mice lacking B cells and immunoglobulins. In contrast, PL-induced inflammation was abrogated in MyD88-deficient mice. PL-induced responses required alveolar macrophages.
Conclusions: These results strongly suggest that PL-induced lung inflammation is dependent on an innate MyD88-dependent pathway rather than the Ig-binding properties of this microbial B cell superantigen. We propose that this pulmonary inflammatory reaction is caused by the interaction of PL with a Toll-like receptor expressed on alveolar macrophages.
Figures
References
-
- Levinson AI, Kozlowski L, Zheng Y, Wheatley L. B-cell superantigens: definition and potential impact on the immune response. J Clin Immunol. 1995;15:26S–36S. - PubMed
-
- Silverman GJ. B-cell superantigens. Immunol Today. 1997;18:379–386. - PubMed
-
- Kozlowski LM, Silverman G, Lambris JD, Levinson AI. Complement activation by a B cell superantigen. J Immunol. 1996;157:1200–12006. 1996. - PubMed
-
- Kozlowski LM, Li W, Goldschmidt M, Levinson AI. In vivo inflammatory response to a prototypic B cell superantigen: elicitation of an Arthus reaction by staphylococcal protein A. J Immunol. 1998;160:5246–5252. 1998. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
