

Functional significance of the taper of vertebrate cone photoreceptors
- PMID: 22250013
- PMCID: PMC3269789
- DOI: 10.1085/jgp.201110692
Functional significance of the taper of vertebrate cone photoreceptors
Abstract
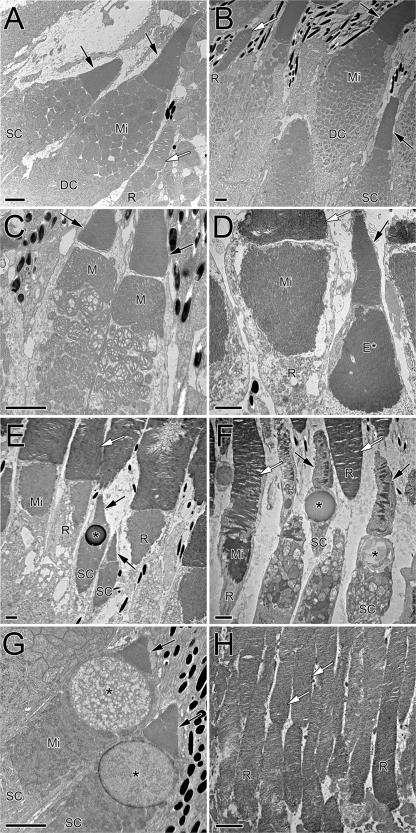
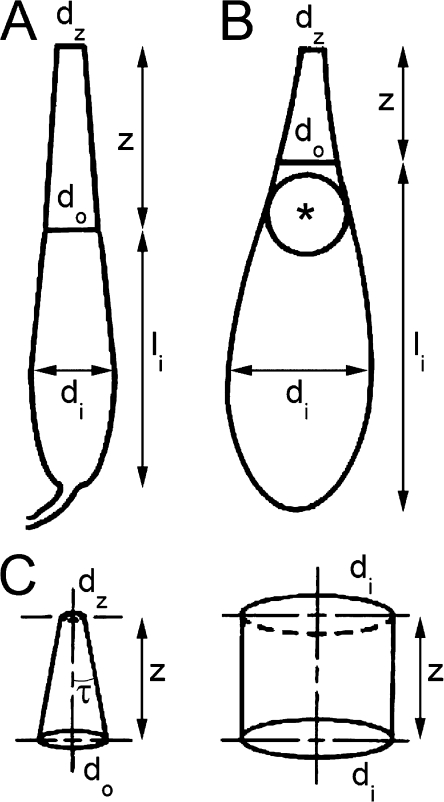
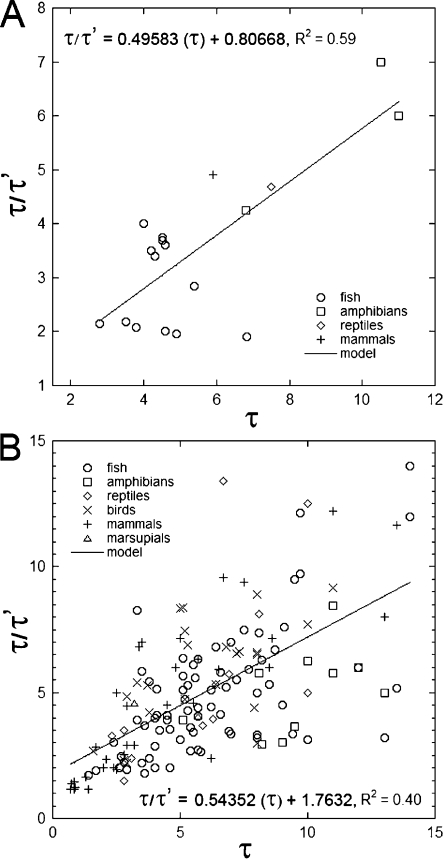
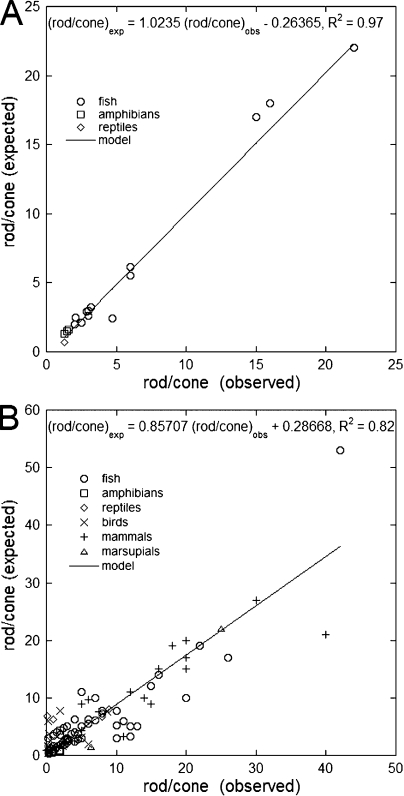
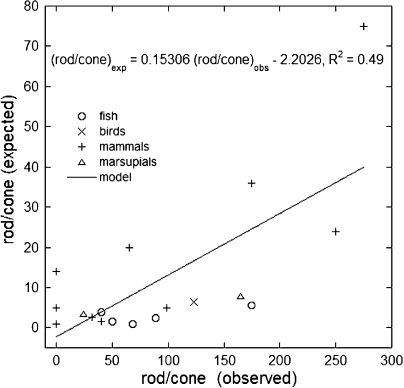
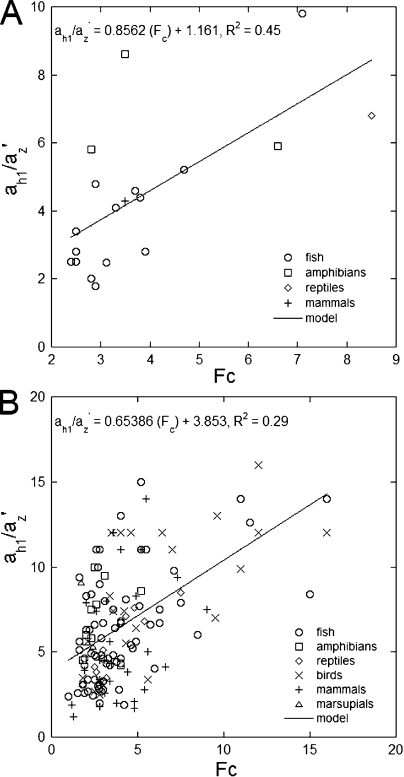
Vertebrate photoreceptors are commonly distinguished based on the shape of their outer segments: those of cones taper, whereas the ones from rods do not. The functional advantages of cone taper, a common occurrence in vertebrate retinas, remain elusive. In this study, we investigate this topic using theoretical analyses aimed at revealing structure-function relationships in photoreceptors. Geometrical optics combined with spectrophotometric and morphological data are used to support the analyses and to test predictions. Three functions are considered for correlations between taper and functionality. The first function proposes that outer segment taper serves to compensate for self-screening of the visual pigment contained within. The second function links outer segment taper to compensation for a signal-to-noise ratio decline along the longitudinal dimension. Both functions are supported by the data: real cones taper more than required for these compensatory roles. The third function relates outer segment taper to the optical properties of the inner compartment whereby the primary determinant is the inner segment's ability to concentrate light via its ellipsoid. In support of this idea, the rod/cone ratios of primarily diurnal animals are predicted based on a principle of equal light flux gathering between photoreceptors. In addition, ellipsoid concentration factor, a measure of ellipsoid ability to concentrate light onto the outer segment, correlates positively with outer segment taper expressed as a ratio of characteristic lengths, where critical taper is the yardstick. Depending on a light-funneling property and the presence of focusing organelles such as oil droplets, cone outer segments can be reduced in size to various degrees. We conclude that outer segment taper is but one component of a miniaturization process that reduces metabolic costs while improving signal detection. Compromise solutions in the various retinas and retinal regions occur between ellipsoid size and acuity, on the one hand, and faster response time and reduced light sensitivity, on the other.
Figures
Similar articles
-
Tuning outer segment Ca2+ homeostasis to phototransduction in rods and cones.Adv Exp Med Biol. 2002;514:179-203. doi: 10.1007/978-1-4615-0121-3_11. Adv Exp Med Biol. 2002. PMID: 12596922 Review.
-
Phosphorylation-independent suppression of light-activated visual pigment by arrestin in carp rods and cones.J Biol Chem. 2015 Apr 10;290(15):9399-411. doi: 10.1074/jbc.M114.634543. Epub 2015 Feb 20. J Biol Chem. 2015. PMID: 25713141 Free PMC article.
-
The retina visual cycle is driven by cis retinol oxidation in the outer segments of cones.Vis Neurosci. 2017 Jan;34:E004. doi: 10.1017/S0952523817000013. Vis Neurosci. 2017. PMID: 28359344 Free PMC article.
-
Photoreceptors of Nrl -/- mice coexpress functional S- and M-cone opsins having distinct inactivation mechanisms.J Gen Physiol. 2005 Mar;125(3):287-304. doi: 10.1085/jgp.200409208. J Gen Physiol. 2005. PMID: 15738050 Free PMC article.
-
Evolution, Development and Function of Vertebrate Cone Oil Droplets.Front Neural Circuits. 2017 Dec 8;11:97. doi: 10.3389/fncir.2017.00097. eCollection 2017. Front Neural Circuits. 2017. PMID: 29276475 Free PMC article. Review.
Cited by
-
Swimming behaviour tunes fish polarization vision to double prey sighting distance.Sci Rep. 2019 Jan 30;9(1):944. doi: 10.1038/s41598-018-37632-1. Sci Rep. 2019. PMID: 30700806 Free PMC article.
-
Single cones give rise to multi-cone types in the retinas of fishes.Sci Rep. 2025 Mar 6;15(1):7823. doi: 10.1038/s41598-025-91987-w. Sci Rep. 2025. PMID: 40050341 Free PMC article.
-
Straying from the flatfish retinal plan: Cone photoreceptor patterning in the common sole (Solea solea) and the Senegalese sole (Solea senegalensis).J Comp Neurol. 2020 Oct;528(14):2283-2307. doi: 10.1002/cne.24893. Epub 2020 Mar 4. J Comp Neurol. 2020. PMID: 32103501 Free PMC article.
-
Evolutionary transformation of rod photoreceptors in the all-cone retina of a diurnal garter snake.Proc Natl Acad Sci U S A. 2016 Jan 12;113(2):356-61. doi: 10.1073/pnas.1513284113. Epub 2015 Dec 29. Proc Natl Acad Sci U S A. 2016. PMID: 26715746 Free PMC article.
-
Optics of cone photoreceptors in the chicken (Gallus gallus domesticus).J R Soc Interface. 2015 Oct 6;12(111):20150591. doi: 10.1098/rsif.2015.0591. J R Soc Interface. 2015. PMID: 26423439 Free PMC article.
References
-
- Ahlbert I.B. 1973. Ontogeny of double cones in the retina of perch fry (Perca fluviatilis, Teleostei). Acta Zoologica (Stockholm, Sweden). 54:241–254 10.1111/j.1463-6395.1973.tb00460.x - DOI
-
- Ali M.A., Anctil M. 1976. Retinas of Fishes: An Atlas. Springer-Verlag, New York. 248 pp.
-
- Ali M.A., Anctil M. 1977. Retinal structure and function in the walleye (Stizostedion vitreum vitreum) and sauger (S. canadense). Journal of the Fisheries Research Board of Canada. 34:1467–1474 10.1139/f77-211 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
