

A quantitative high-throughput in vitro splicing assay identifies inhibitors of spliceosome catalysis
- PMID: 22252314
- PMCID: PMC3302438
- DOI: 10.1128/MCB.05788-11
A quantitative high-throughput in vitro splicing assay identifies inhibitors of spliceosome catalysis
Abstract
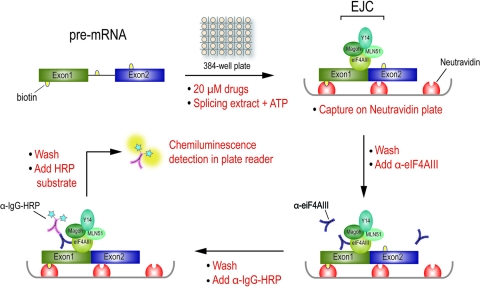
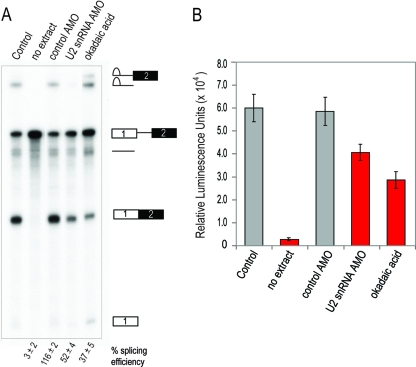
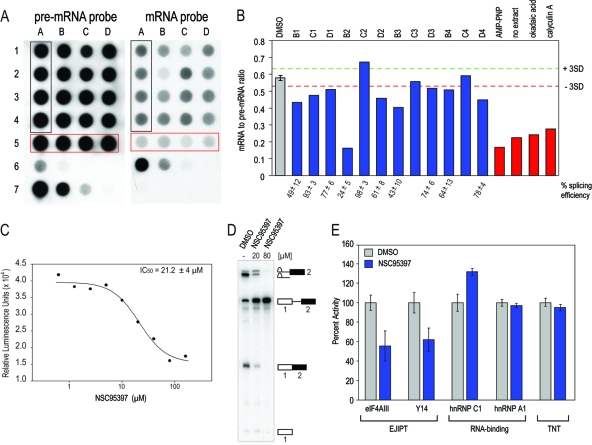
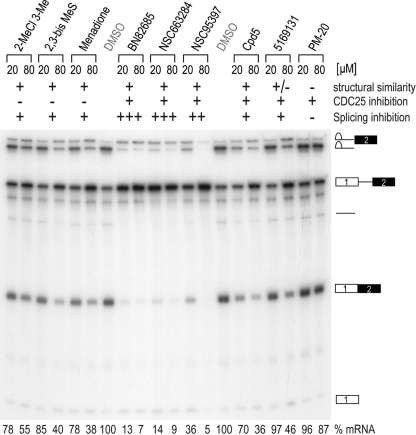
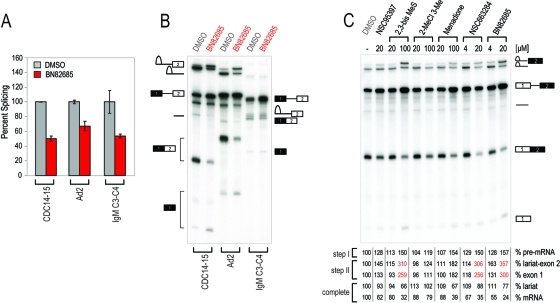
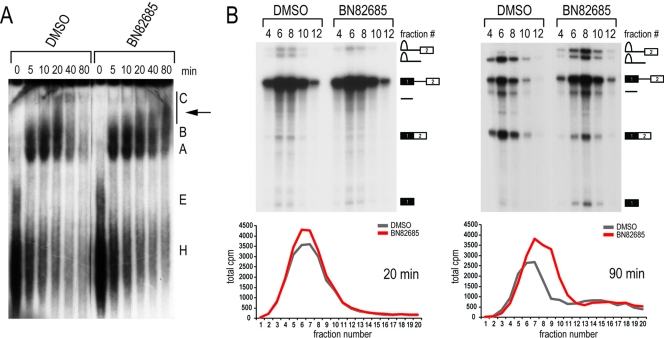
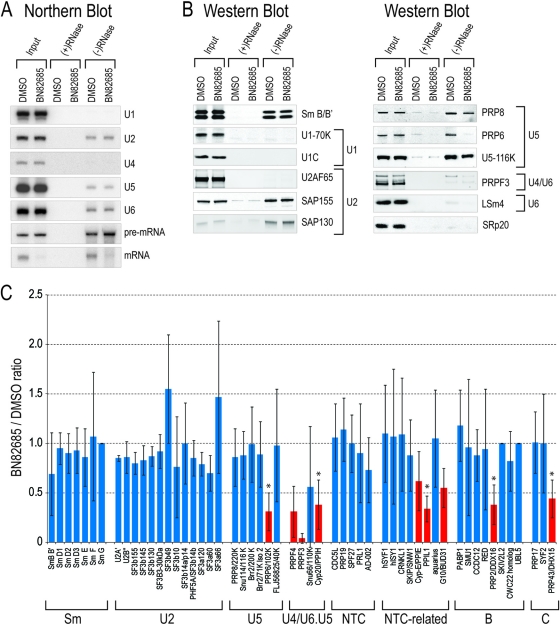
Despite intensive research, there are very few reagents with which to modulate and dissect the mRNA splicing pathway. Here, we describe a novel approach to identify such tools, based on detection of the exon junction complex (EJC), a unique molecular signature that splicing leaves on mRNAs. We developed a high-throughput, splicing-dependent EJC immunoprecipitation (EJIPT) assay to quantitate mRNAs spliced from biotin-tagged pre-mRNAs in cell extracts, using antibodies to EJC components Y14 and eukaryotic translation initiation factor 4aIII (eIF4AIII). Deploying EJIPT we performed high-throughput screening (HTS) in conjunction with secondary assays to identify splicing inhibitors. We describe the identification of 1,4-naphthoquinones and 1,4-heterocyclic quinones with known anticancer activity as potent and selective splicing inhibitors. Interestingly, and unlike previously described small molecules, most of which target early steps, our inhibitors represented by the benzothiazole-4,7-dione, BN82685, block the second of two trans-esterification reactions in splicing, preventing the release of intron lariat and ligation of exons. We show that BN82685 inhibits activated spliceosomes' elaborate structural rearrangements that are required for second-step catalysis, allowing definition of spliceosomes stalled in midcatalysis. EJIPT provides a platform for characterization and discovery of splicing and EJC modulators.
Figures
Similar articles
-
A high-throughput splicing assay identifies new classes of inhibitors of human and yeast spliceosomes.J Biomol Screen. 2013 Oct;18(9):1110-20. doi: 10.1177/1087057113493117. Epub 2013 Jun 14. J Biomol Screen. 2013. PMID: 23771823 Free PMC article.
-
CWC22-dependent pre-mRNA splicing and eIF4A3 binding enables global deposition of exon junction complexes.Nucleic Acids Res. 2015 May 19;43(9):4687-700. doi: 10.1093/nar/gkv320. Epub 2015 Apr 13. Nucleic Acids Res. 2015. PMID: 25870412 Free PMC article.
-
Human spliceosomal protein CWC22 plays a role in coupling splicing to exon junction complex deposition and nonsense-mediated decay.Proc Natl Acad Sci U S A. 2012 Dec 26;109(52):21313-8. doi: 10.1073/pnas.1219725110. Epub 2012 Dec 10. Proc Natl Acad Sci U S A. 2012. PMID: 23236153 Free PMC article.
-
Molecular choreography of pre-mRNA splicing by the spliceosome.Curr Opin Struct Biol. 2019 Dec;59:124-133. doi: 10.1016/j.sbi.2019.07.010. Epub 2019 Aug 30. Curr Opin Struct Biol. 2019. PMID: 31476650 Review.
-
The spliceosome and its metal ions.Met Ions Life Sci. 2011;9:235-51. doi: 10.1039/9781849732512-00235. Met Ions Life Sci. 2011. PMID: 22010274 Review.
Cited by
-
Telomere dysfunction drives aberrant hematopoietic differentiation and myelodysplastic syndrome.Cancer Cell. 2015 May 11;27(5):644-57. doi: 10.1016/j.ccell.2015.04.007. Cancer Cell. 2015. PMID: 25965571 Free PMC article.
-
Identification of Genomewide Alternative Splicing Events in Sequential, Isogenic Clinical Isolates of Candida albicans Reveals a Novel Mechanism of Drug Resistance and Tolerance to Cellular Stresses.mSphere. 2020 Aug 12;5(4):e00608-20. doi: 10.1128/mSphere.00608-20. mSphere. 2020. PMID: 32817456 Free PMC article.
-
A high-throughput splicing assay identifies new classes of inhibitors of human and yeast spliceosomes.J Biomol Screen. 2013 Oct;18(9):1110-20. doi: 10.1177/1087057113493117. Epub 2013 Jun 14. J Biomol Screen. 2013. PMID: 23771823 Free PMC article.
-
Identification of small molecule inhibitors of pre-mRNA splicing.J Biol Chem. 2014 Dec 12;289(50):34683-98. doi: 10.1074/jbc.M114.590976. Epub 2014 Oct 3. J Biol Chem. 2014. PMID: 25281741 Free PMC article.
-
Small Molecule Modulators of Pre-mRNA Splicing in Cancer Therapy.Trends Mol Med. 2016 Jan;22(1):28-37. doi: 10.1016/j.molmed.2015.11.005. Epub 2015 Dec 14. Trends Mol Med. 2016. PMID: 26700537 Free PMC article. Review.
References
-
- Achsel T, Ahrens K, Brahms H, Teigelkamp S, Luhrmann R. 1998. The human U5-220kD protein (hPrp8) forms a stable RNA-free complex with several U5-specific proteins, including an RNA unwindase, a homologue of ribosomal elongation factor EF-2, and a novel WD-40 protein. Mol. Cell. Biol. 18:6756–6766 - PMC - PubMed
-
- Ballut L, et al. 2005. The exon junction core complex is locked onto RNA by inhibition of eIF4AIII ATPase activity. Nat. Struct. Mol. Biol. 12:861–869 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
