

XIer2 is required for convergent extension movements during Xenopus development
- PMID: 22252488
- PMCID: PMC3261581
- DOI: 10.1387/ijdb.113288sh
XIer2 is required for convergent extension movements during Xenopus development
Abstract
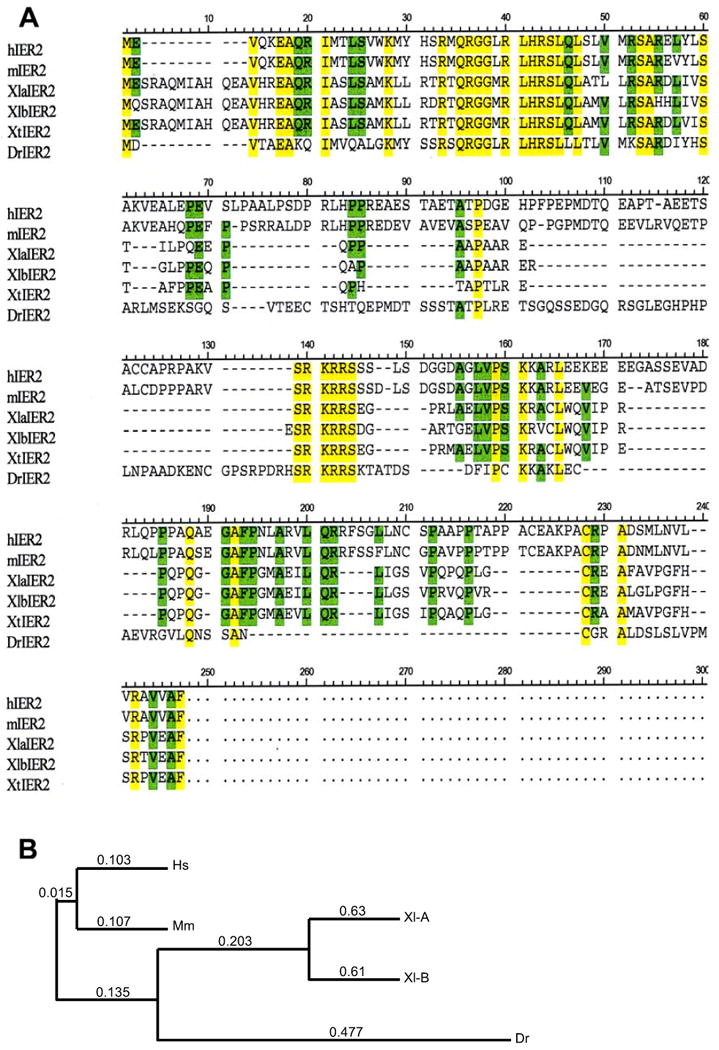
Immediate early response 2 (Ier2) is a downstream target of fibroblast growth factor (FGF) signaling. In zebrafish, Ier2 is involved in left-right asymmetry establishment and in convergent extension movements. We isolated the Xenopus ier2 gene based on sequence similarity searches using multiple vertebrate species. Xenopus Ier2 has high homology in the N-terminal region to other vertebrate Ier2 proteins, and Xier2 transcripts were observed from oocytes through larval stages. Except for the maternal expression of xier2, the expression of this gene in the marginal region at gastrulation and in somites and the notochord at later stages is similar to the expression pattern of zebrafish ier2. XIer2 knockdown using antisense morpholinos resulted in defects of convergent extension leading to severe neural tube defects; overexpression of Ier2 showed similar, albeit milder phenotypes. Assays in animal cap explants likewise showed inhibition of elongation after blocking XIer2 expression. These results indicate that Xenopus Ier2 is essential for the execution of convergent extension movements during early Xenopus development.
Figures



Similar articles
-
The planar polarity gene strabismus regulates convergent extension movements in Xenopus.EMBO J. 2002 Mar 1;21(5):976-85. doi: 10.1093/emboj/21.5.976. EMBO J. 2002. PMID: 11867525 Free PMC article.
-
FGF-dependent left-right asymmetry patterning in zebrafish is mediated by Ier2 and Fibp1.Proc Natl Acad Sci U S A. 2009 Feb 17;106(7):2230-5. doi: 10.1073/pnas.0812880106. Epub 2009 Jan 21. Proc Natl Acad Sci U S A. 2009. PMID: 19164561 Free PMC article.
-
A novel gene, BENI is required for the convergent extension during Xenopus laevis gastrulation.Dev Biol. 2007 Mar 1;303(1):270-80. doi: 10.1016/j.ydbio.2006.11.014. Epub 2006 Nov 11. Dev Biol. 2007. PMID: 17174295
-
Convergent extension in the amphibian, Xenopus laevis.Curr Top Dev Biol. 2020;136:271-317. doi: 10.1016/bs.ctdb.2019.11.013. Epub 2019 Dec 27. Curr Top Dev Biol. 2020. PMID: 31959291 Free PMC article. Review.
-
The use of antisense oligonucleotides in Xenopus oocytes.Methods. 2010 May;51(1):75-81. doi: 10.1016/j.ymeth.2009.12.015. Epub 2010 Jan 5. Methods. 2010. PMID: 20045732 Free PMC article. Review.
Cited by
-
A zebrafish embryo screen utilizing gastrulation identifies the HTR2C inhibitor pizotifen as a suppressor of EMT-mediated metastasis.Elife. 2021 Dec 17;10:e70151. doi: 10.7554/eLife.70151. Elife. 2021. PMID: 34919051 Free PMC article.
-
Gastrulation Screening to Identify Anti-metastasis Drugs in Zebrafish Embryos.Bio Protoc. 2022 Oct 5;12(19):e4525. doi: 10.21769/BioProtoc.4525. eCollection 2022 Oct 5. Bio Protoc. 2022. PMID: 36313195 Free PMC article.
-
Full transcriptome analysis of early dorsoventral patterning in zebrafish.PLoS One. 2013 Jul 29;8(7):e70053. doi: 10.1371/journal.pone.0070053. Print 2013. PLoS One. 2013. PMID: 23922899 Free PMC article.
References
-
- Chung HA, Hyodo-Miura J, Nagamune T, Ueno N. FGF signal regulates gastrulation cell movements and morphology through its target NRH. Dev Biol. 2005;282:95–110. - PubMed
-
- Conlon FL, Smith JC. Interference with brachyury function inhibits convergent extension, causes apoptosis, and reveals separate requirements in the FGF and activin signalling pathways. Dev Biol. 1999;213:85–100. - PubMed
-
- Harland RM. In situ hybridization: an improved whole-mount method for Xenopus embryos. Methods Cell Biol. 1991;36:685–695. - PubMed
-
- Heisenberg CP, Tada M, Rauch GJ, Saúde L, Concha ML, Geisler R, Stemple DL, Smith JC, Wilson SW. Silberblick/Wnt11 mediates convergent extension movements during zebrafish gastrulation. Nature. 2000;405:76–81. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
