

Leaf-induced gibberellin signaling is essential for internode elongation, cambial activity, and fiber differentiation in tobacco stems
- PMID: 22253226
- PMCID: PMC3289570
- DOI: 10.1105/tpc.111.093096
Leaf-induced gibberellin signaling is essential for internode elongation, cambial activity, and fiber differentiation in tobacco stems
Abstract
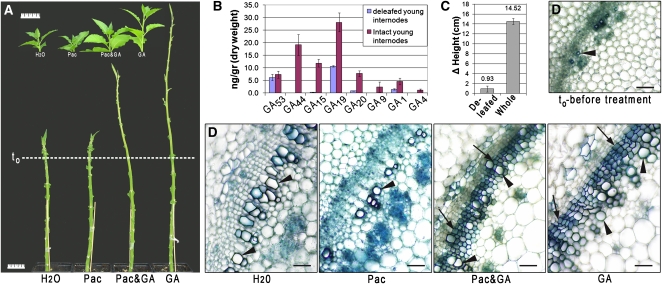
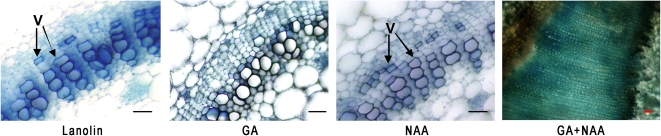
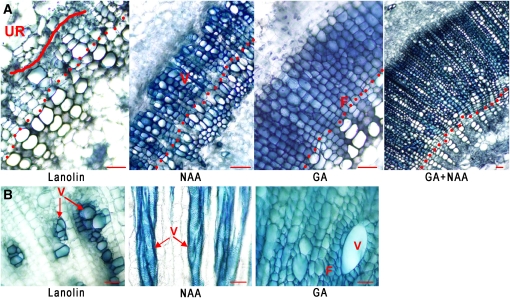
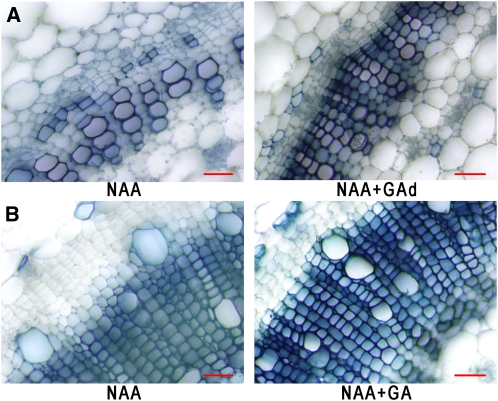
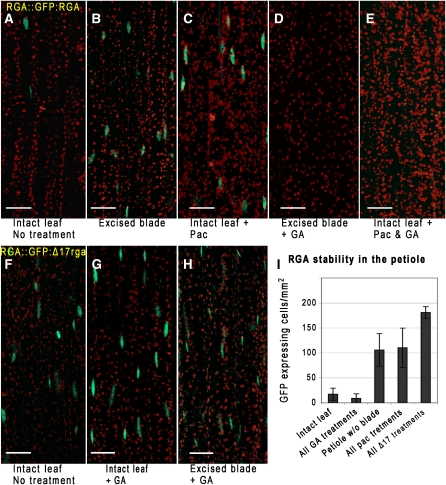
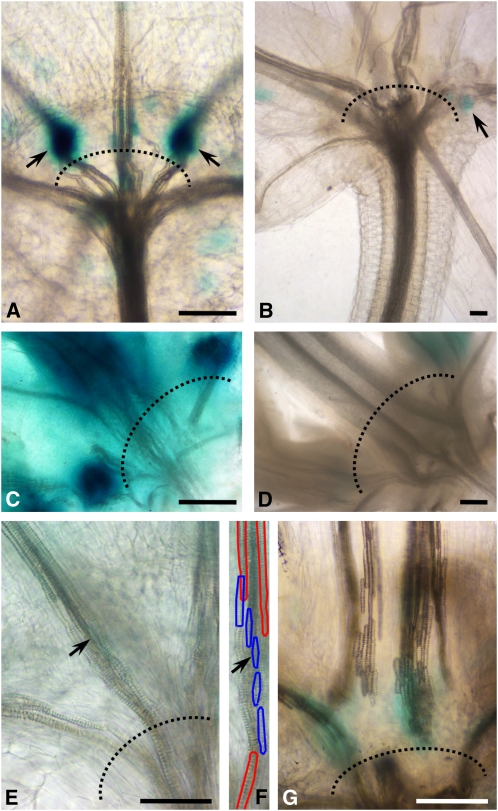
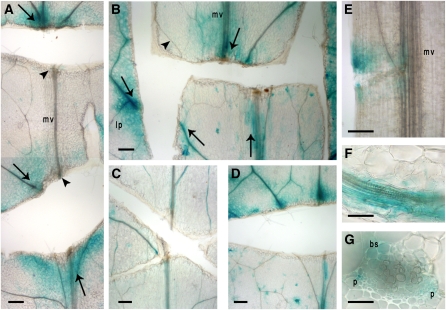
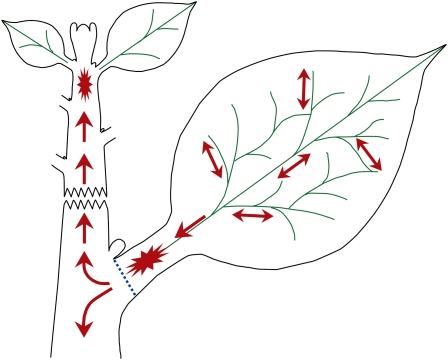
The gibberellins (GAs) are a group of endogenous compounds that promote the growth of most plant organs, including stem internodes. We show that in tobacco (Nicotiana tabacum) the presence of leaves is essential for the accumulation of bioactive GAs and their immediate precursors in the stem and consequently for normal stem elongation, cambial proliferation, and xylem fiber differentiation. These processes do not occur in the absence of maturing leaves but can be restored by application of C(19)-GAs, identifying the presence of leaves as a requirement for GA signaling in stems and revealing the fundamental role of GAs in secondary growth regulation. The use of reporter genes for GA activity and GA-directed DELLA protein degradation in Arabidopsis thaliana confirms the presence of a mobile signal from leaves to the stem that induces GA signaling.
Figures


References
-
- Achard P., Cheng H., De Grauwe L., Decat J., Schoutteten H., Moritz T., Van Der Straeten D., Peng J., Harberd N.P. (2006). Integration of plant responses to environmentally activated phytohormonal signals. Science 311: 91–94 - PubMed
-
- Aloni R. (1987). Differentiation of vascular tissues. Annu. Rev. Plant Physiol. 38: 179–204
-
- Aloni R. (2010). The induction of vascular tissues by auxin. In Plant Hormones: Biosynthesis, Signal Transduction, Action! P.J. Davies, ed (Dordrecht, The Netherlands: Kluwer Academic), pp. 471–492
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
