

A new type of compartment, defined by plant-specific Atg8-interacting proteins, is induced upon exposure of Arabidopsis plants to carbon starvation
- PMID: 22253227
- PMCID: PMC3289568
- DOI: 10.1105/tpc.111.093112
A new type of compartment, defined by plant-specific Atg8-interacting proteins, is induced upon exposure of Arabidopsis plants to carbon starvation
Abstract
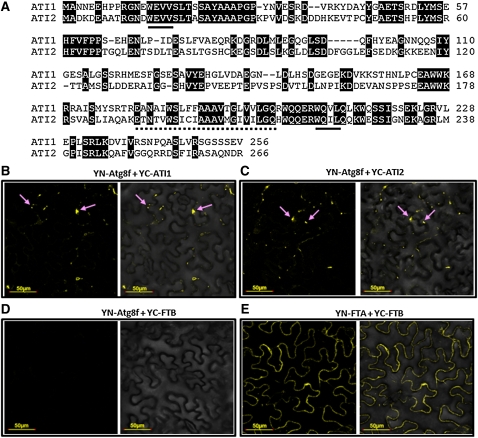
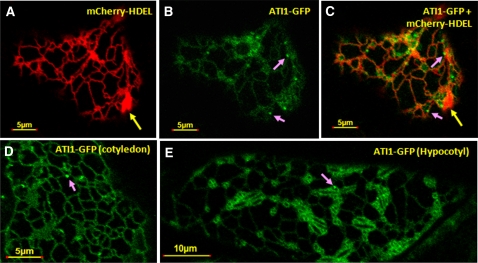
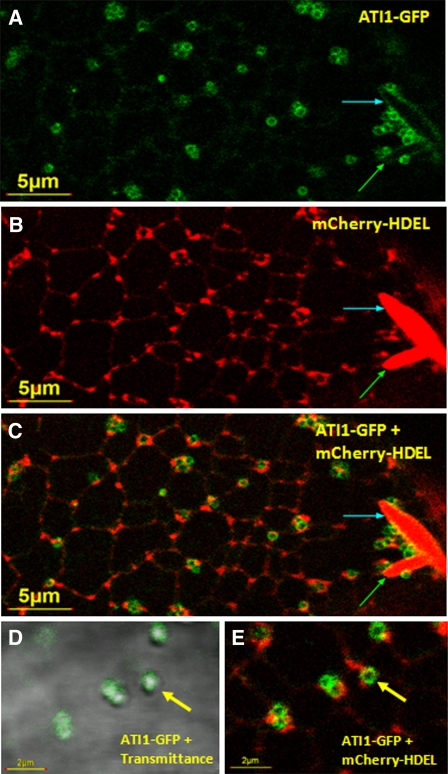
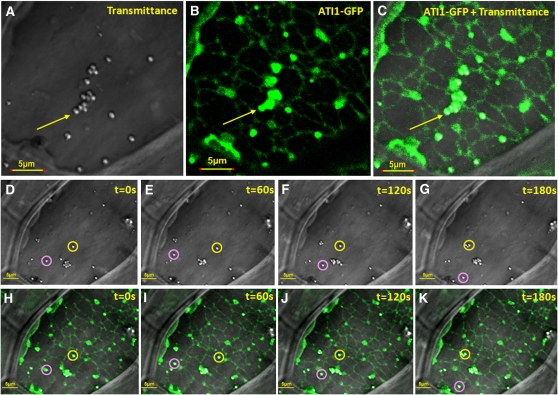
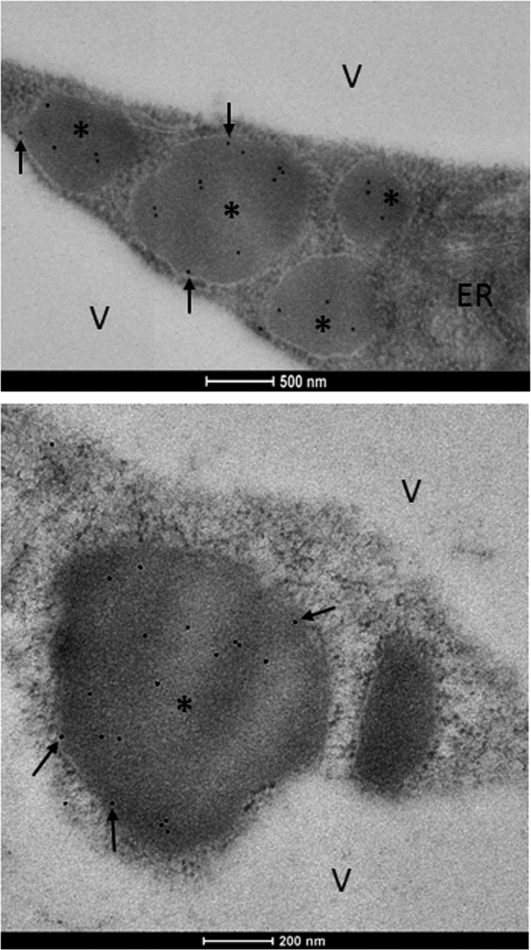
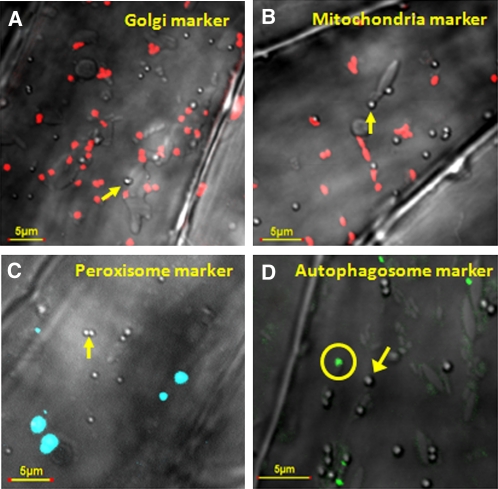
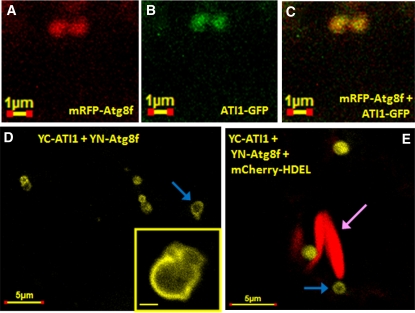
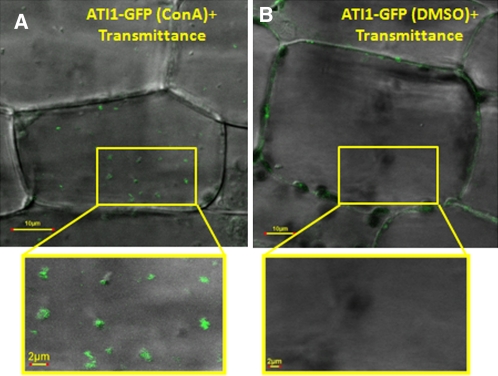
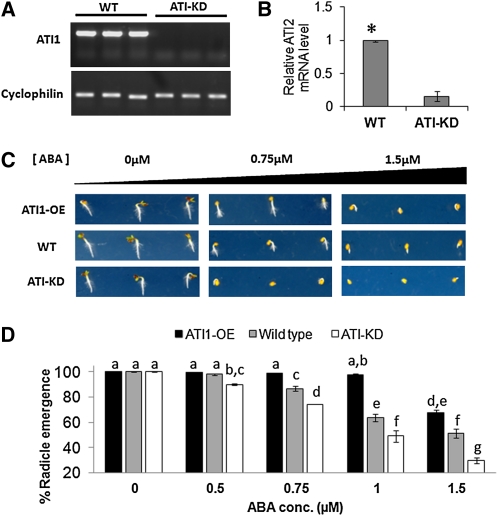
Atg8 is a central protein in bulk starvation-induced autophagy, but it is also specifically associated with multiple protein targets under various physiological conditions to regulate their selective turnover by the autophagy machinery. Here, we describe two new closely related Arabidopsis thaliana Atg8-interacting proteins (ATI1 and ATI2) that are unique to plants. We show that under favorable growth conditions, ATI1 and ATI2 are partially associated with the endoplasmic reticulum (ER) membrane network, whereas upon exposure to carbon starvation, they become mainly associated with newly identified spherical compartments that dynamically move along the ER network. These compartments are morphologically distinct from previously reported spindle-shaped ER bodies and, in contrast to them, do not contain ER-lumenal markers possessing a C-terminal HDEL sequence. Organelle and autophagosome-specific markers show that the bodies containing ATI1 are distinct from Golgi, mitochondria, peroxisomes, and classical autophagosomes. The final destination of the ATI1 bodies is the central vacuole, indicating that they may operate in selective turnover of specific proteins. ATI1 and ATI2 gene expression is elevated during late seed maturation and desiccation. We further demonstrate that ATI1 overexpression or suppression of both ATI1 and ATI2, respectively, stimulate or inhibit seed germination in the presence of the germination-inhibiting hormone abscisic acid.
Figures
Similar articles
-
ATI1 (ATG8-interacting protein 1) and ATI2 define a plant starvation-induced reticulophagy pathway and serve as MSBP1/MAPR5 cargo receptors.Autophagy. 2021 Nov;17(11):3375-3388. doi: 10.1080/15548627.2021.1872886. Epub 2021 Jan 25. Autophagy. 2021. PMID: 33487099 Free PMC article.
-
Arabidopsis ATG8-INTERACTING PROTEIN1 is involved in autophagy-dependent vesicular trafficking of plastid proteins to the vacuole.Plant Cell. 2014 Oct;26(10):4084-101. doi: 10.1105/tpc.114.129999. Epub 2014 Oct 3. Plant Cell. 2014. PMID: 25281689 Free PMC article.
-
The transmembrane autophagy cargo receptors ATI1 and ATI2 interact with ATG8 through intrinsically disordered regions with distinct biophysical properties.Biochem J. 2019 Feb 5;476(3):449-465. doi: 10.1042/BCJ20180748. Biochem J. 2019. PMID: 30642888
-
Genes for plant autophagy: functions and interactions.Mol Cells. 2012 Nov;34(5):413-23. doi: 10.1007/s10059-012-0098-y. Epub 2012 Jul 6. Mol Cells. 2012. PMID: 22772908 Free PMC article. Review.
-
Involvement of autophagy in the direct ER to vacuole protein trafficking route in plants.Front Plant Sci. 2014 Apr 8;5:134. doi: 10.3389/fpls.2014.00134. eCollection 2014. Front Plant Sci. 2014. PMID: 24782875 Free PMC article. Review.
Cited by
-
Heterologous expression of ATG8c from soybean confers tolerance to nitrogen deficiency and increases yield in Arabidopsis.PLoS One. 2012;7(5):e37217. doi: 10.1371/journal.pone.0037217. Epub 2012 May 22. PLoS One. 2012. PMID: 22629371 Free PMC article.
-
Marchantia polymorpha, a New Model Plant for Autophagy Studies.Front Plant Sci. 2019 Jul 17;10:935. doi: 10.3389/fpls.2019.00935. eCollection 2019. Front Plant Sci. 2019. PMID: 31379911 Free PMC article.
-
hfAIM: A reliable bioinformatics approach for in silico genome-wide identification of autophagy-associated Atg8-interacting motifs in various organisms.Autophagy. 2016 May 3;12(5):876-87. doi: 10.1080/15548627.2016.1147668. Epub 2016 Apr 12. Autophagy. 2016. PMID: 27071037 Free PMC article.
-
Emerging Roles of the Selective Autophagy in Plant Immunity and Stress Tolerance.Int J Mol Sci. 2020 Aug 31;21(17):6321. doi: 10.3390/ijms21176321. Int J Mol Sci. 2020. PMID: 32878263 Free PMC article. Review.
-
ROOT HAIR DEFECTIVE3 Is a Receptor for Selective Autophagy of the Endoplasmic Reticulum in Arabidopsis.Front Plant Sci. 2022 Feb 24;13:817251. doi: 10.3389/fpls.2022.817251. eCollection 2022. Front Plant Sci. 2022. PMID: 35283874 Free PMC article.
References
-
- Alonso J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Bassham D.C. (2007). Plant autophagy—More than a starvation response. Curr. Opin. Plant Biol. 10: 587–593 - PubMed
-
- Bassham D.C. (2009). Function and regulation of macroautophagy in plants. Biochim. Biophys. Acta 1793: 1397–1403 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
