

Dynamic ligand modulation of EPO receptor pools, and dysregulation by polycythemia-associated EPOR alleles
- PMID: 22253704
- PMCID: PMC3257245
- DOI: 10.1371/journal.pone.0029064
Dynamic ligand modulation of EPO receptor pools, and dysregulation by polycythemia-associated EPOR alleles
Abstract
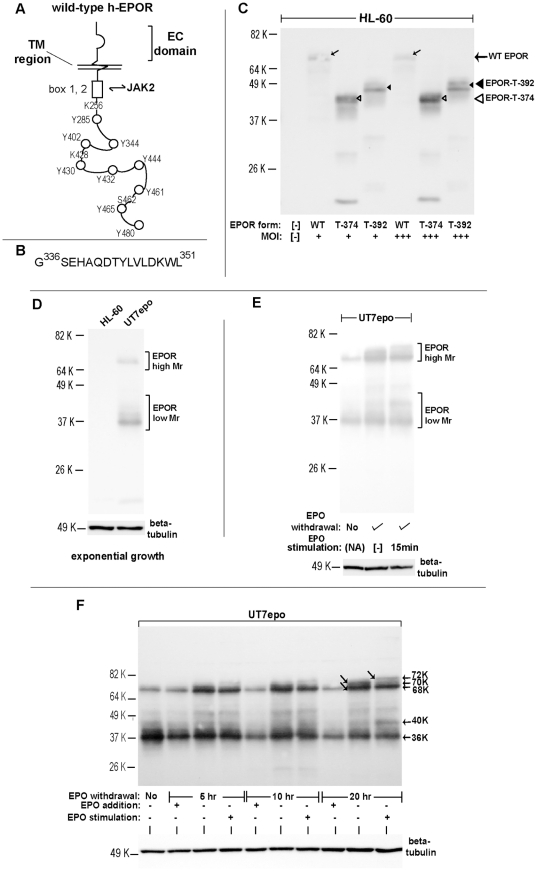
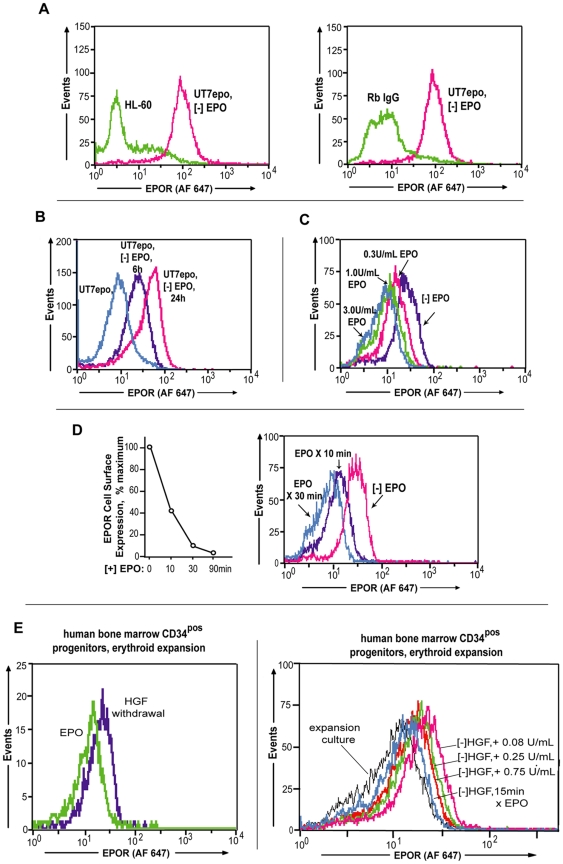
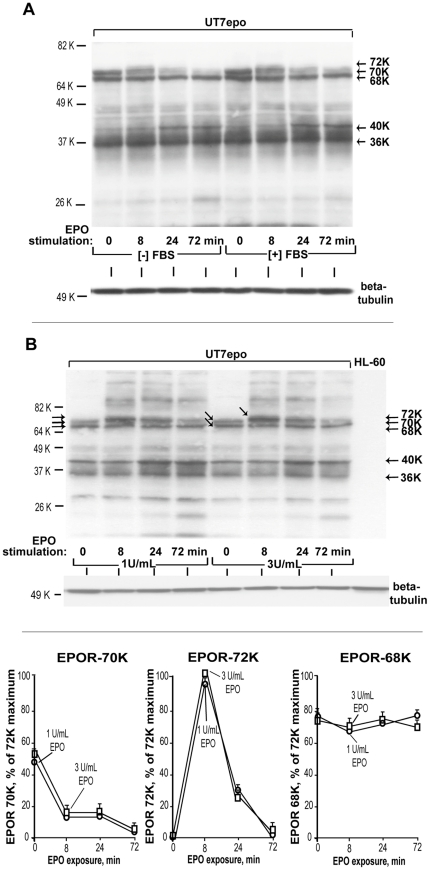
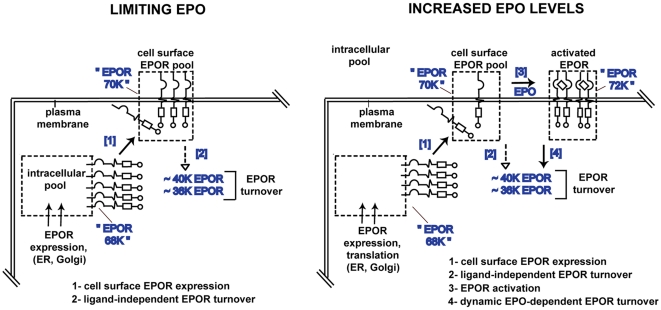
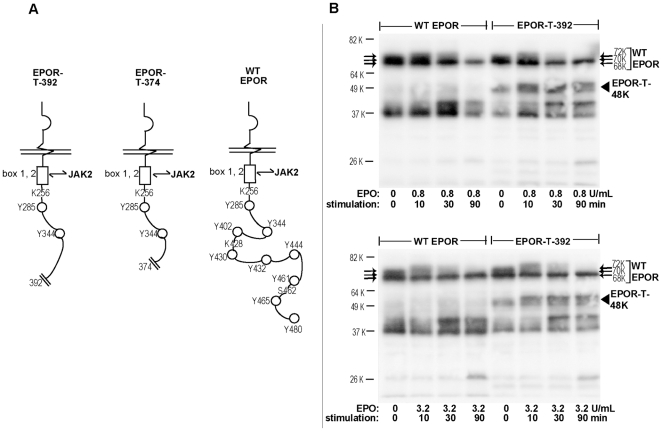
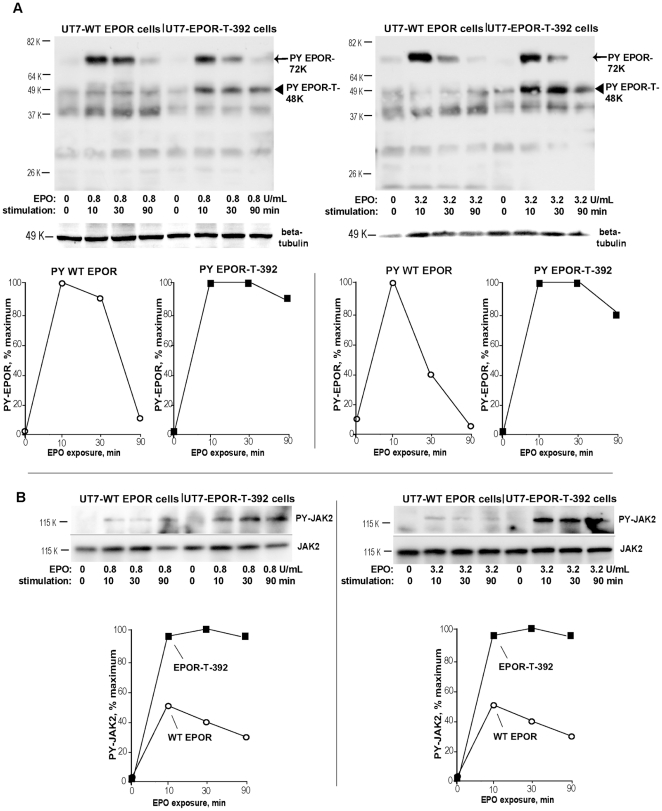
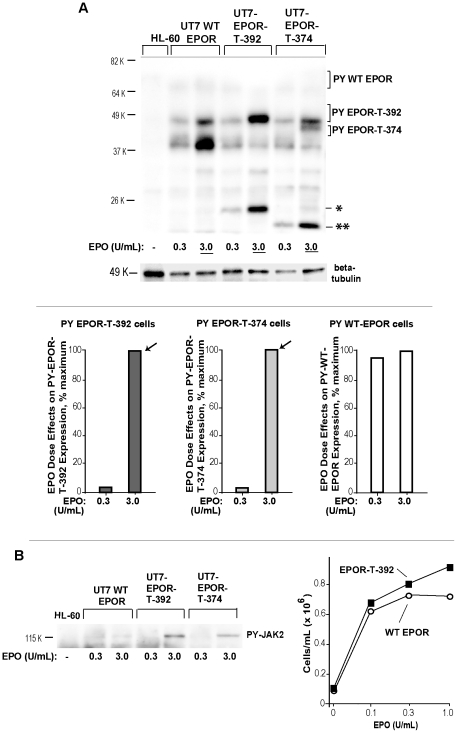
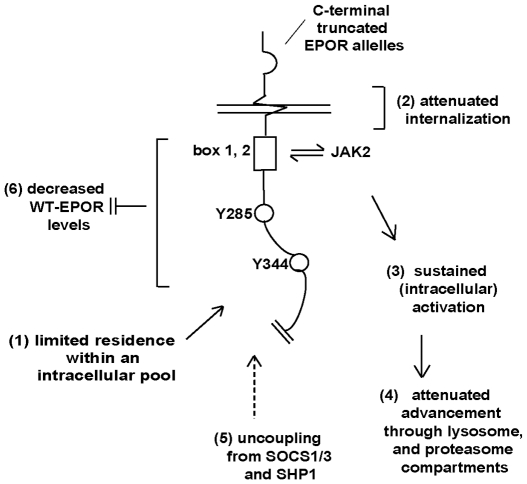
Erythropoietin (EPO) and its cell surface receptor (EPOR) are essential for erythropoiesis; can modulate non-erythroid target tissues; and have been reported to affect the progression of certain cancers. Basic studies of EPOR expression and trafficking, however, have been hindered by low-level EPOR occurrence, and the limited specificity of anti-EPOR antibodies. Consequently, these aspects of EPOR biology are not well defined, nor are actions of polycythemia- associated mutated EPOR alleles. Using novel rabbit monoclonal antibodies to intracellular, PY- activated and extracellular EPOR domains, the following properties of the endogenous hEPOR in erythroid progenitors first are unambiguously defined. 1) High- Mr EPOR forms become obviously expressed only when EPO is limited. 2) EPOR-68K plus -70K species sequentially accumulate, and EPOR-70K comprises an apparent cell surface EPOR population. 3) Brefeldin A, N-glycanase and associated analyses point to EPOR-68K as a core-glycosylated intracellular EPOR pool (of modest size). 4) In contrast to recent reports, EPOR inward trafficking is shown (in UT7epo cells, and primary proerythroblasts) to be sharply ligand-dependent. Beyond this, when C-terminal truncated hEPOR-T mutant alleles as harbored by polycythemia patients are co-expressed with the wild-type EPOR in EPO-dependent erythroid progenitors, several specific events become altered. First, EPOR-T alleles are persistently activated upon EPO- challenge, yet are also subject to apparent turn-over (to low-Mr EPOR products). Furthermore, during exponential cell growth EPOR-T species become both over-represented, and hyper-activated. Interestingly, EPOR-T expression also results in an EPO dose-dependent loss of endogenous wild-type EPOR's (and, therefore, a squelching of EPOR C-terminal- mediated negative feedback effects). New knowledge concerning regulated EPOR expression and trafficking therefore is provided, together with new insight into mechanisms via which mutated EPOR-T polycythemia alleles dysregulate the erythron. Notably, specific new tools also are characterized for studies of EPOR expression, activation, action and metabolism.
Conflict of interest statement
Figures


Similar articles
-
EPO receptor gain-of-function causes hereditary polycythemia, alters CD34 cell differentiation and increases circulating endothelial precursors.PLoS One. 2010 Aug 5;5(8):e12015. doi: 10.1371/journal.pone.0012015. PLoS One. 2010. Retraction in: PLoS One. 2020 Mar 4;15(3):e0230279. doi: 10.1371/journal.pone.0230279. PMID: 20700488 Free PMC article. Retracted.
-
Erythropoietin hypersensitivity in primary familial and congenital polycythemia: role of tyrosines Y285 and Y344 in erythropoietin receptor cytoplasmic domain.Biochim Biophys Acta. 2005 Apr 15;1740(1):17-28. doi: 10.1016/j.bbadis.2005.03.003. Epub 2005 Apr 1. Biochim Biophys Acta. 2005. PMID: 15878737
-
Primary familial polycythemia: a frameshift mutation in the erythropoietin receptor gene and increased sensitivity of erythroid progenitors to erythropoietin.Blood. 1995 Jul 1;86(1):15-22. Blood. 1995. PMID: 7795221
-
The human erythropoietin receptor.Int J Cell Cloning. 1992 Sep;10(5):254-61. doi: 10.1002/stem.5530100502. Int J Cell Cloning. 1992. PMID: 1453011 Review.
-
"Benign erythrocytosis" and other familial and congenital polycythemias.Eur J Haematol. 1996 Oct;57(4):263-8. Eur J Haematol. 1996. PMID: 8982288 Review.
Cited by
-
RHEX, a novel regulator of human erythroid progenitor cell expansion and erythroblast development.J Exp Med. 2014 Aug 25;211(9):1715-22. doi: 10.1084/jem.20130624. Epub 2014 Aug 4. J Exp Med. 2014. PMID: 25092874 Free PMC article.
-
Governing roles for Trib3 pseudokinase during stress erythropoiesis.Exp Hematol. 2017 May;49:48-55.e5. doi: 10.1016/j.exphem.2016.12.010. Epub 2017 Jan 4. Exp Hematol. 2017. PMID: 28062363 Free PMC article.
-
Phospho-PTM proteomic discovery of novel EPO- modulated kinases and phosphatases, including PTPN18 as a positive regulator of EPOR/JAK2 Signaling.Cell Signal. 2020 May;69:109554. doi: 10.1016/j.cellsig.2020.109554. Epub 2020 Feb 3. Cell Signal. 2020. PMID: 32027948 Free PMC article.
-
The hypoxia inducible factor/erythropoietin (EPO)/EPO receptor pathway is disturbed in a rat model of chronic kidney disease related anemia.PLoS One. 2018 May 8;13(5):e0196684. doi: 10.1371/journal.pone.0196684. eCollection 2018. PLoS One. 2018. PMID: 29738538 Free PMC article.
-
Emerging EPO and EPO receptor regulators and signal transducers.Blood. 2015 Jun 4;125(23):3536-41. doi: 10.1182/blood-2014-11-575357. Epub 2015 Apr 17. Blood. 2015. PMID: 25887776 Free PMC article. Review.
References
-
- Perugini M, Varelias A, Sadlon T, D'Andrea RJ. Hematopoietic growth factor mimetics: from concept to clinic. Cytokine Growth Factor Rev. 2009;20:87–94. - PubMed
-
- Baker SJ, Rane SG, Reddy EP. Hematopoietic cytokine receptor signaling. Oncogene. 2007;26:6724–6737. - PubMed
-
- Pedersen M, Ronnstrand L, Sun J. The c-Kit/D816V mutation eliminates the differences in signal transduction and biological responses between two isoforms of c-Kit. Cell Signal. 2009;21:413–418. - PubMed
-
- Chen J, Olsen J, Ford S, Mirza S, Walker A, et al. A new isoform of interleukin-3 receptor {alpha} with novel differentiation activity and high affinity binding mode. J Biol Chem. 2009;284:5763–5773. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
