

Spatial modeling of vesicle transport and the cytoskeleton: the challenge of hitting the right road
- PMID: 22253752
- PMCID: PMC3257240
- DOI: 10.1371/journal.pone.0029645
Spatial modeling of vesicle transport and the cytoskeleton: the challenge of hitting the right road
Abstract
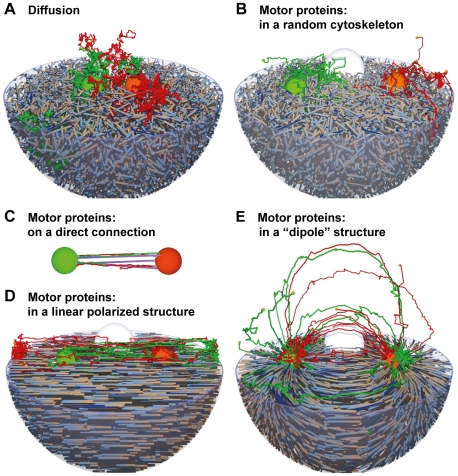
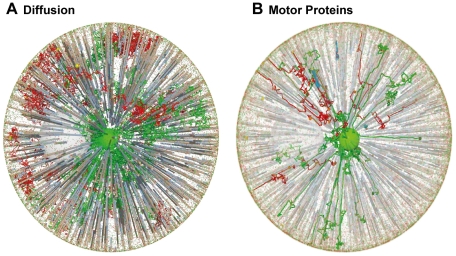
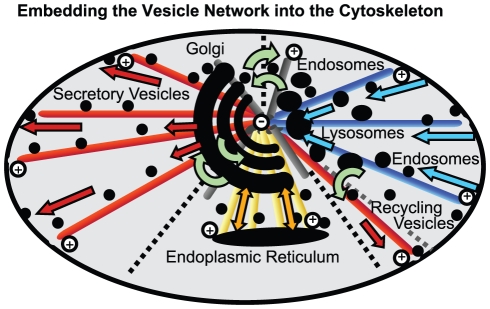
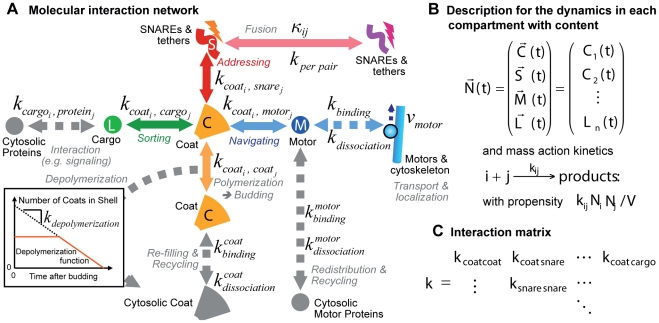
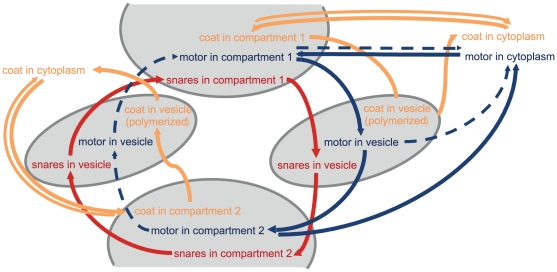
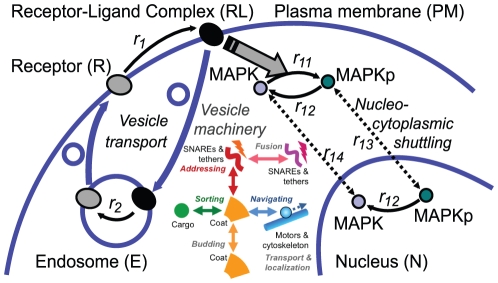
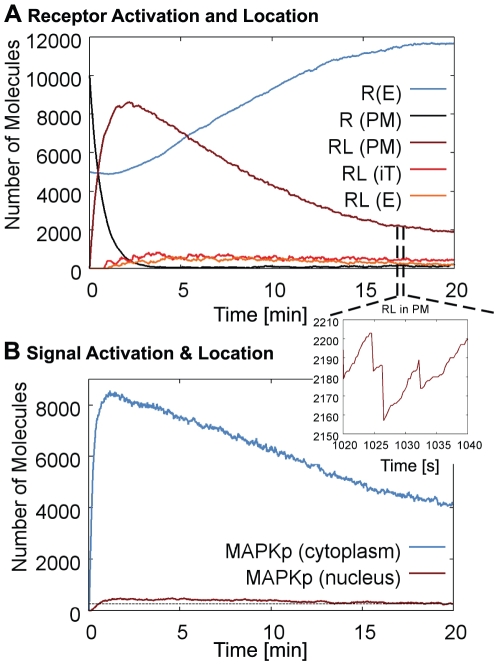
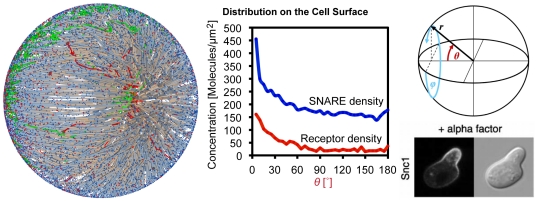
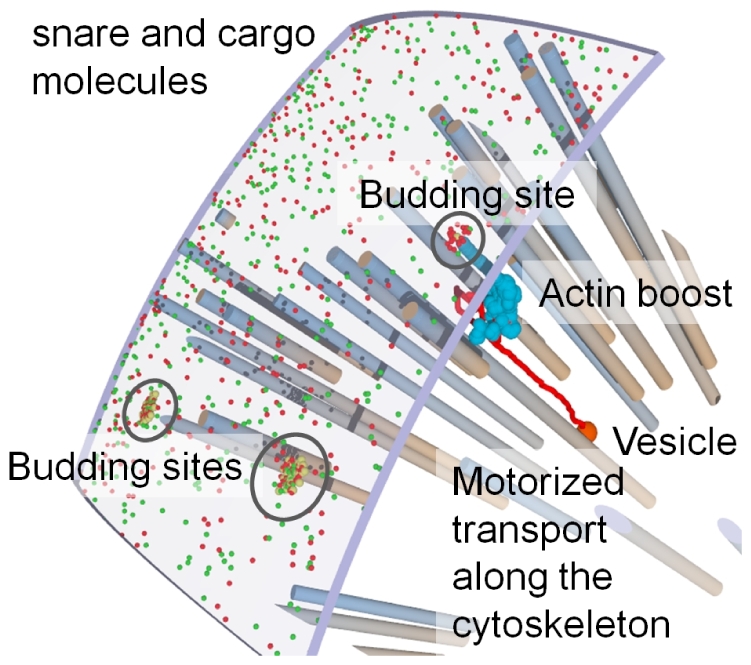
The membrane trafficking machinery provides a transport and sorting system for many cellular proteins. We propose a mechanistic agent-based computer simulation to integrate and test the hypothesis of vesicle transport embedded into a detailed model cell. The method tracks both the number and location of the vesicles. Thus both the stochastic properties due to the low numbers and the spatial aspects are preserved. The underlying molecular interactions that control the vesicle actions are included in a multi-scale manner based on the model of Heinrich and Rapoport (2005). By adding motor proteins we can improve the recycling process of SNAREs and model cell polarization. Our model also predicts that coat molecules should have a high turnover at the compartment membranes, while the turnover of motor proteins has to be slow. The modular structure of the underlying model keeps it tractable despite the overall complexity of the vesicle system. We apply our model to receptor-mediated endocytosis and show how a polarized cytoskeleton structure leads to polarized distributions in the plasma membrane both of SNAREs and the Ste2p receptor in yeast. In addition, we can couple signal transduction and membrane trafficking steps in one simulation, which enables analyzing the effect of receptor-mediated endocytosis on signaling.
Conflict of interest statement
Figures
Similar articles
-
The syndapin protein family: linking membrane trafficking with the cytoskeleton.J Cell Sci. 2004 Jul 1;117(Pt 15):3077-86. doi: 10.1242/jcs.01290. J Cell Sci. 2004. PMID: 15226389 Review.
-
Calcium-Sensing Receptor: Trafficking, Endocytosis, Recycling, and Importance of Interacting Proteins.Prog Mol Biol Transl Sci. 2015;132:127-50. doi: 10.1016/bs.pmbts.2015.02.006. Epub 2015 Apr 25. Prog Mol Biol Transl Sci. 2015. PMID: 26055057 Review.
-
Cytoplasmic protein motility and polarized sorting during asymmetric cell division.Wiley Interdiscip Rev Dev Biol. 2013 Nov-Dec;2(6):797-808. doi: 10.1002/wdev.116. Epub 2013 Apr 1. Wiley Interdiscip Rev Dev Biol. 2013. PMID: 24123938 Review.
-
Trafficking of the plasma membrane gamma-aminobutyric acid transporter GAT1. Size and rates of an acutely recycling pool.J Biol Chem. 2005 May 13;280(19):18703-9. doi: 10.1074/jbc.M500381200. Epub 2005 Mar 18. J Biol Chem. 2005. PMID: 15778221
-
Regulation of membrane traffic by integrin signaling.Trends Cell Biol. 2011 May;21(5):266-73. doi: 10.1016/j.tcb.2011.02.003. Epub 2011 Mar 26. Trends Cell Biol. 2011. PMID: 21440440 Review.
Cited by
-
A multiscale model of the regulation of aquaporin 2 recycling.NPJ Syst Biol Appl. 2022 May 9;8(1):16. doi: 10.1038/s41540-022-00223-y. NPJ Syst Biol Appl. 2022. PMID: 35534498 Free PMC article.
-
Spatial simulations in systems biology: from molecules to cells.Int J Mol Sci. 2012;13(6):7798-7827. doi: 10.3390/ijms13067798. Epub 2012 Jun 21. Int J Mol Sci. 2012. PMID: 22837728 Free PMC article. Review.
-
Spatial redistribution of neurosecretory vesicles upon stimulation accelerates their directed transport to the plasma membrane.PLoS One. 2022 Mar 16;17(3):e0264521. doi: 10.1371/journal.pone.0264521. eCollection 2022. PLoS One. 2022. PMID: 35294476 Free PMC article.
-
Real time measures of prestin charge and fluorescence during plasma membrane trafficking reveal sub-tetrameric activity.PLoS One. 2013 Jun 10;8(6):e66078. doi: 10.1371/journal.pone.0066078. Print 2013. PLoS One. 2013. PMID: 23762468 Free PMC article.
-
Involvement of small G proteins and vesicle traffic in the glutoxim and molixan effects on the intracellular Ca(2+) concentration in macrophages.Dokl Biol Sci. 2014 Jul;457(1):252-4. doi: 10.1134/S0012496614040036. Epub 2014 Aug 30. Dokl Biol Sci. 2014. PMID: 25172594 No abstract available.
References
-
- Olkkonen V, Ikonen E. When intracellular logistics fails-genetic defects in membrane traf- ficking. Journal of Cell Science. 2006;119:5031. - PubMed
-
- Derby M, Gleeson P. New insights into membrane trafficking and protein sorting. Int Rev Cytol. 2007;261:47–116. - PubMed
-
- Bonifacino J, Glick B. The mechanisms of vesicle budding and fusion. Cell. 2004;116:153–166. - PubMed
-
- Bowers K, Stevens T. Protein transport from the late Golgi to the vacuole in the yeast Saccharomyces cerevisiae. BBA-Molecular Cell Research. 2005;1744:438–454. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
