

Müller glial cell-provided cellular light guidance through the vital guinea-pig retina
- PMID: 22261048
- PMCID: PMC3297812
- DOI: 10.1016/j.bpj.2011.09.062
Müller glial cell-provided cellular light guidance through the vital guinea-pig retina
Abstract
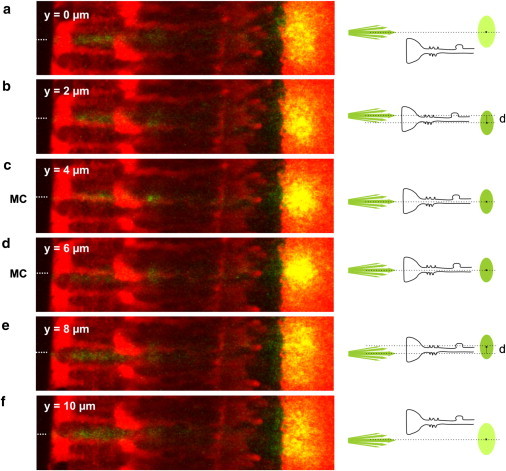
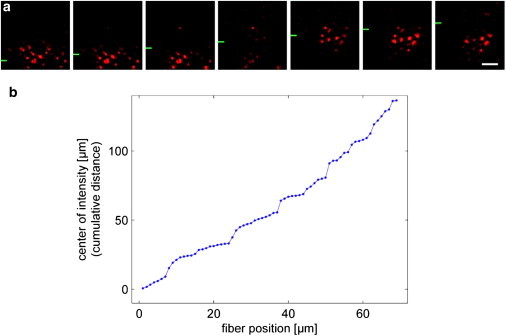
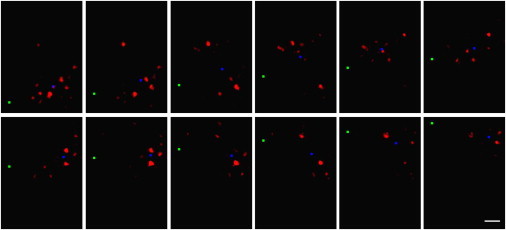
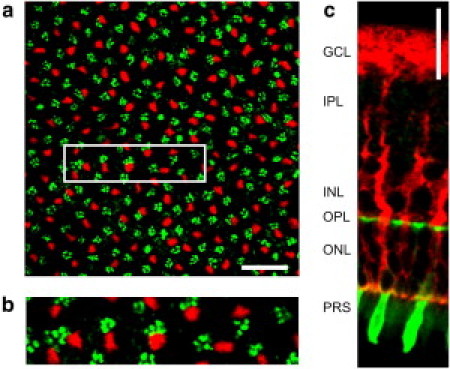
In vertebrate eyes, images are projected onto an inverted retina where light passes all retinal layers on its way to the photoreceptor cells. Light scattering within this tissue should impair vision. We show that radial glial (Müller) cells in the living retina minimize intraretinal light scatter and conserve the diameter of a beam that hits a single Müller cell endfoot. Thus, light arrives at individual photoreceptors with high intensity. This leads to an optimized signal/noise ratio, which increases visual sensitivity and contrast. Moreover, we show that the ratio between Müller cells and cones-responsible for acute vision-is roughly 1. This suggests that high spatiotemporal resolution may be achieved by each cone receiving its part of the image via its individual Müller cell-light guide.
Copyright © 2011 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
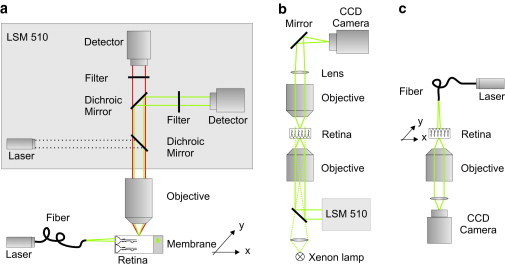
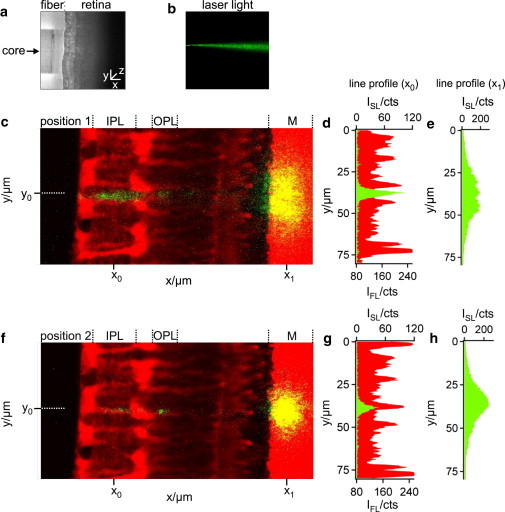
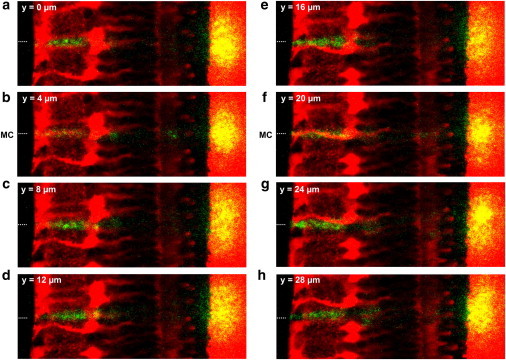
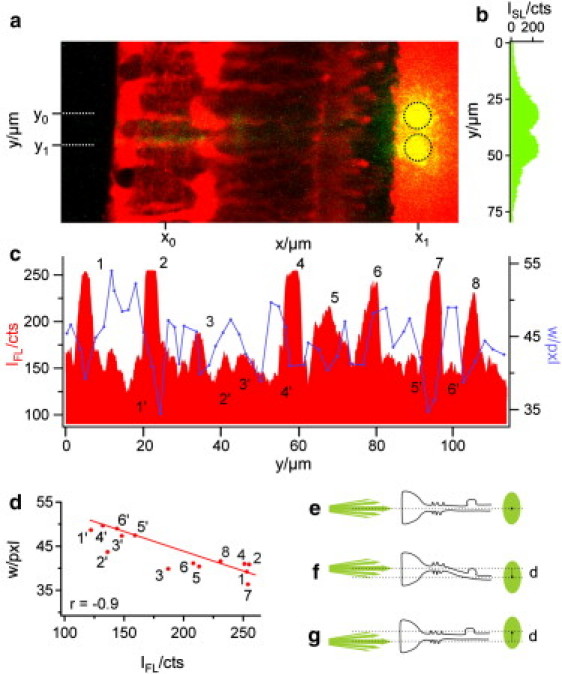

References
-
- Kröger R.H., Biehlmaier O. Space-saving advantage of an inverted retina. Vision Res. 2009;49:2318–2321. - PubMed
-
- Nguyen-Legros J., Hicks D. Renewal of photoreceptor outer segments and their phagocytosis by the retinal pigment epithelium. Int. Rev. Cytol. 2000;196:245–313. - PubMed
-
- Brudenall D.K., Schwab I.R., Graydon M.L. Optimized architecture for nutrition in the avascular retina of Megachiroptera. Anat. Histol. Embryol. 2007;36:382–388. - PubMed
-
- Goldsmith T.H. Optimization, constraint, and history in the evolution of eyes. Q. Rev. Biol. 1990;65:281–322. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
