

Apolipoprotein C-I binds more strongly to phospholipid/triolein/water than triolein/water interfaces: a possible model for inhibiting cholesterol ester transfer protein activity and triacylglycerol-rich lipoprotein uptake
- PMID: 22264166
- PMCID: PMC3313556
- DOI: 10.1021/bi2015212
Apolipoprotein C-I binds more strongly to phospholipid/triolein/water than triolein/water interfaces: a possible model for inhibiting cholesterol ester transfer protein activity and triacylglycerol-rich lipoprotein uptake
Abstract
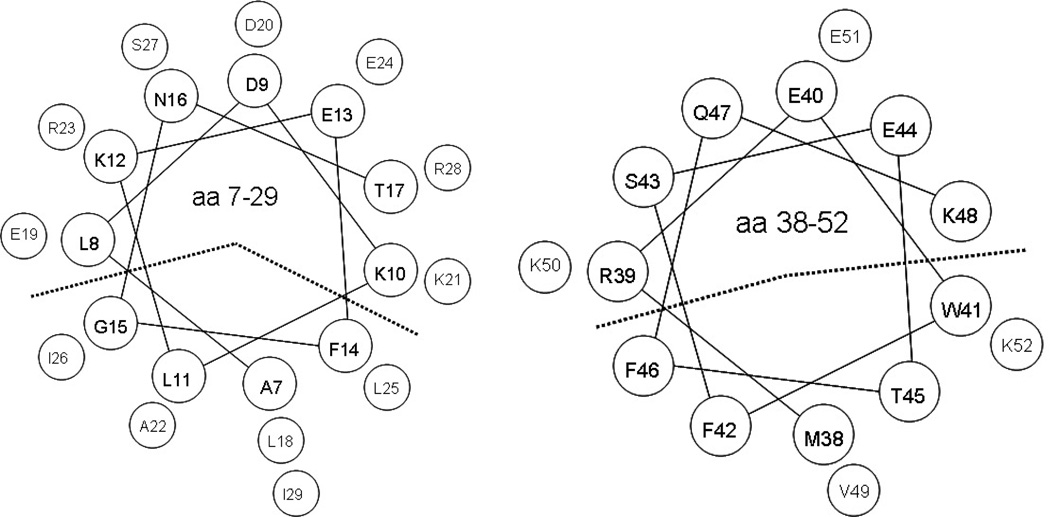
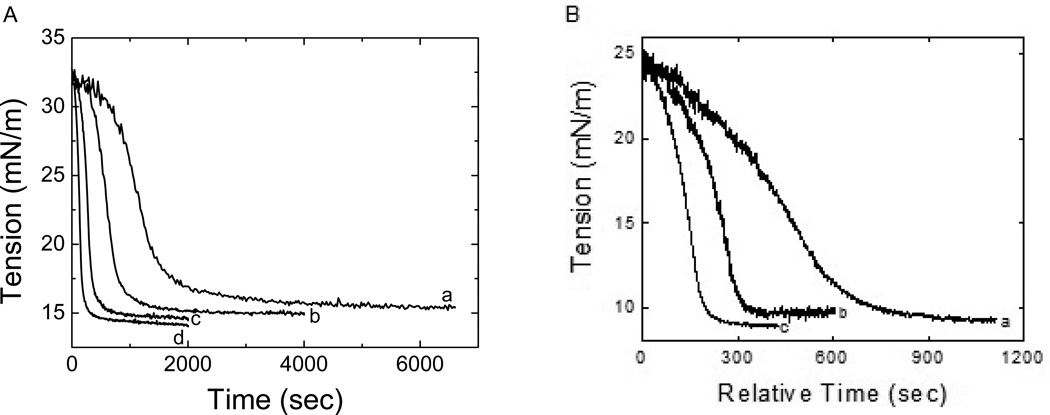
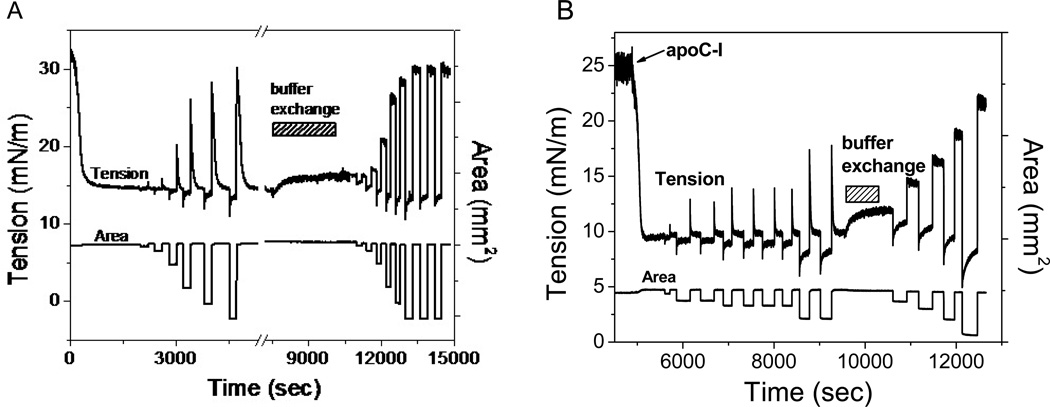
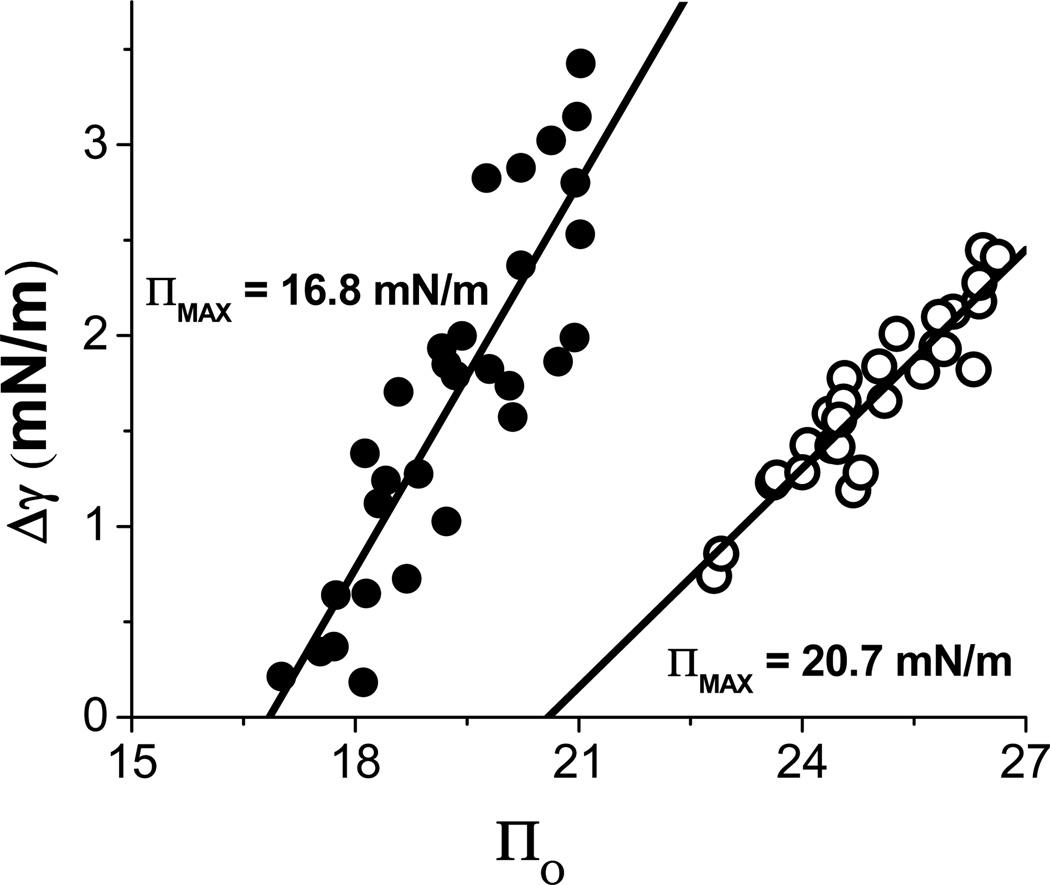
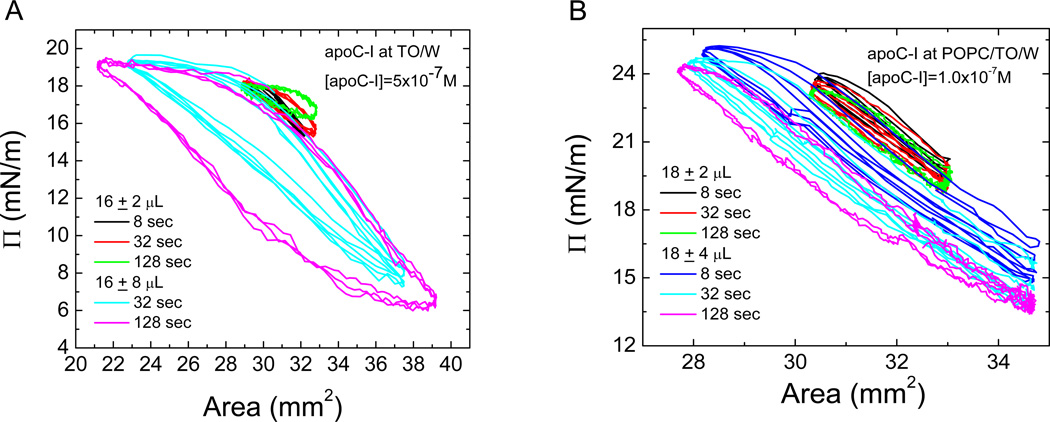
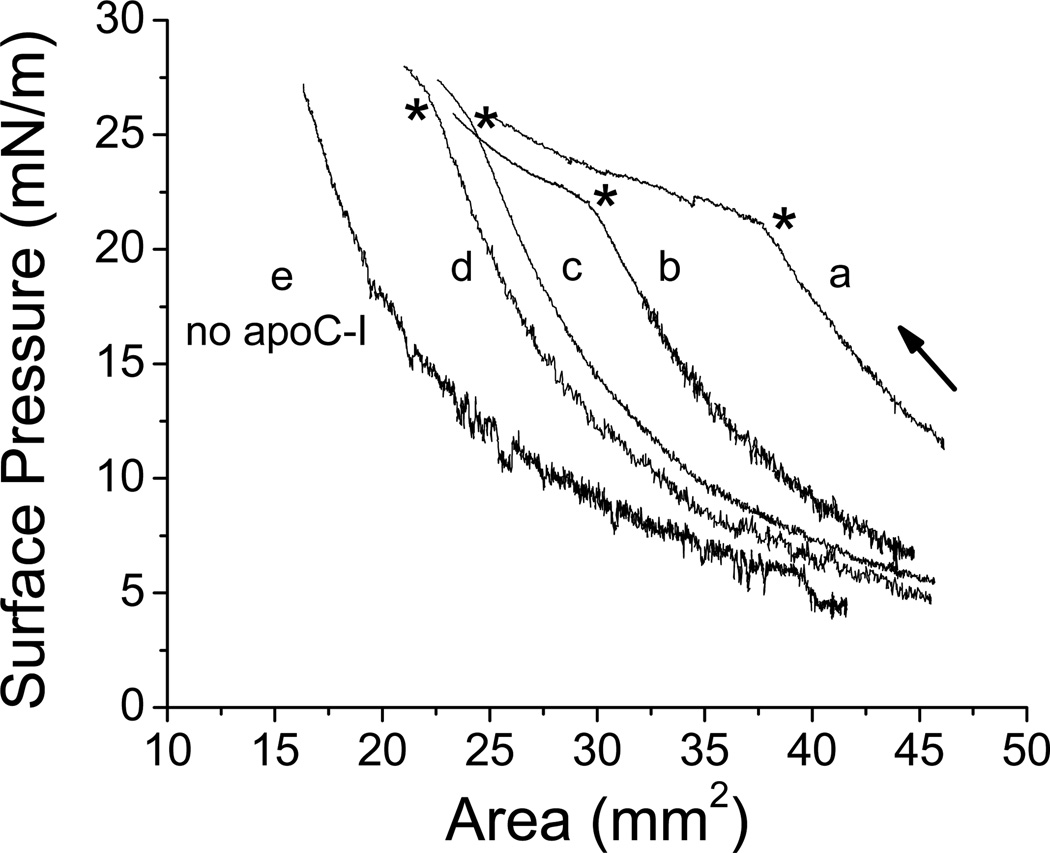
Apolipoprotein C-I (apoC-I) is an important constituent of high-density lipoprotein (HDL) and is involved in the accumulation of cholesterol ester in nascent HDL via inhibition of cholesterol ester transfer protein and potential activation of lecithin:cholesterol acyltransferase (LCAT). As the smallest exchangeable apolipoprotein (57 residues), apoC-I transfers between lipoproteins via a lipid-binding motif of two amphipathic α-helices (AαHs), spanning residues 7-29 and 38-52. To understand apoC-I's behavior at hydrophobic lipoprotein surfaces, oil drop tensiometry was used to compare the binding to triolein/water (TO/W) and palmitoyloleoylphosphatidylcholine/triolein/water (POPC/TO/W) interfaces. When apoC-I binds to either interface, the surface tension (γ) decreases by ~16-18 mN/m. ApoC-I can be exchanged at both interfaces, desorbing upon compression and readsorbing on expansion. The maximal surface pressures at which apoC-I begins to desorb (Π(max)) were 16.8 and 20.7 mN/m at TO/W and POPC/TO/W interfaces, respectively. This suggests that apoC-I interacts with POPC to increase its affinity for the interface. ApoC-I is more elastic on POPC/TO/W than TO/W interfaces, marked by higher values of the elasticity modulus (ε) on oscillations. At POPC/TO/W interfaces containing an increasing POPC:TO ratio, the pressure at which apoC-I begins to be ejected increases as the phospholipid surface concentration increases. The observed increase in apoC-I interface affinity due to higher degrees of apoC-I-POPC interactions may explain how apoC-I can displace larger apolipoproteins, such as apoE, from lipoproteins. These interactions allow apoC-I to remain bound to the interface at higher Π values, offering insight into apoC-I's rearrangement on triacylglycerol-rich lipoproteins as they undergo Π changes during lipoprotein maturation by plasma factors such as lipoprotein lipase.
Figures
References
-
- Schaefer EJ, Eisenberg S, Levy RI. Lipoprotein apoprotein metabolism. J. Lipid Res. 1978;19(6):667–687. - PubMed
-
- Mahley RW, Innerarity TL, Rall SC, Weisgraber KH. Plasma lipoproteins: apolipoprotein structure and function. J. Lipid Res. 1984;25(12):1277–1294. - PubMed
-
- Windler E, Havel RJ. Inhibitory effects of C apolipoproteins from rats and humans on the uptake of triglyceride-rich lipoproteins and their remnants by the perfused rat liver. J. Lipid Res. 1985;26(5):556–565. - PubMed
-
- Soutar AK, Garner CW, Baker HN, Sparrow JT, Jackson RL, Gotto AM, Smith LC. Effect of the human plasma apolipoproteins and phosphatidylcholine acyl donor on the activity of lecithin:cholesterol acyltransferase. Biochemistry. 1975;14(14):3057–3064. - PubMed
-
- Jonas A, Sweeny SA, Herbert PN. Discoidal complexes of A and C apolipoproteins with lipids and their reactions with lecithin:cholesterol acyltransferase. J. Biol. Chem. 1984;259(10):6369–6375. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
