

doi: 10.1016/j.devcel.2011.12.003.
Expanding the morphogenetic repertoire: perspectives from the Drosophila egg
Affiliations
- PMID: 22264728
- PMCID: PMC3266552
- DOI: 10.1016/j.devcel.2011.12.003
Item in Clipboard
Expanding the morphogenetic repertoire: perspectives from the Drosophila egg
Dev Cell.
.
Abstract
Tissue and organ architectures are incredibly diverse, yet our knowledge of the morphogenetic behaviors that generate them is relatively limited. Recent studies have revealed unexpected mechanisms that drive axis elongation in the Drosophila egg, including an unconventional planar polarity signaling pathway, a distinctive type of morphogenetic movement termed "global tissue rotation," a molecular corset-like role of extracellular matrix, and oscillating basal cellular contractions. We review here what is known about Drosophila egg elongation, compare it to other instances of morphogenesis, and highlight several issues of general developmental relevance.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures

(A) Ovariole expressing Collagen-IV-GFP (green), stained for filamentous actin (red), DNA (blue) and FasIII (orange). Stages are marked. Anterior of ovariole is to left and posterior is to right; all subsequent panels share this orientation unless otherwise specified. (B) Close-up of stalk cells (sc) separating two follicles. (C) Anterior-posterior axis of the follicle, established during initial stages of oogenesis, is reflected by polar cells (FasIII-GFP,orange) in the epithelium. The oocyte is localized at the posterior. St. 7 follicle is stained as in (A). (D) Radial symmetry of the follicle at st. 8 demonstrated by slices through the A-P (D’) and circumferential (D”) axes. Indy-GFP (green) marks follicle epithelial plasma membranes.
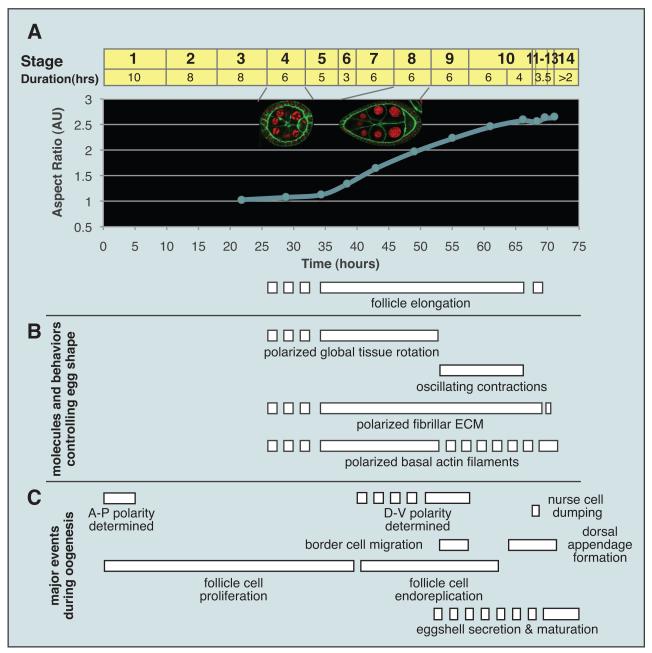
(A) Elongation of follicles, depicted as follicle aspect ratio with respect to developmental time. Stages of oogenesis are marked on top of graph. Insets show st. 4 (isotropic) and st. 8 (elongated) follicles. (B) Timing of events involved in egg elongation, in comparison to major events of oogenesis (C). Dotted lines indicate process initiated but not mature. See also Horne-Badovinac and Bilder (2005).

(A) Planar polarized organization of basal filamentous actin (red) at st. 8. Monopolar protrusions (arrows) reveal coordinated chirality within the tissue. (B) Appearance of planar polarized stress fibres at st. 12. (C) Planar polarized organization of Collagen IV (green) at st. 8. (D) WT elongated egg. (E) Round egg produced by a fat2 homozygous mutant female. (F) Round egg produced by a female with mosaic βPS Integrin mutant clones in the follicle epithelium. (G) Mispolarization of basal actin in fat2 follicle at st. 8. Note signs of local alignment. (H) Aberrant organization of planar polarized actin in βPS Integrin mosaic epithelium at st. 8. Clone boundaries are marked with dotted line; clone is to top. (I) displays same genotype at st. 12. Note non- autonomous polarity defects in neighboring WT cells. (J) Diagram of planar polarized actin and Collagen IV localization before (left) and during (right) follicle elongation. In stage 4 follicles, Collagen IV fibrils are nascent and basal actin polarization is poorly coordinated. After stage 5, Collagen IV fibrils elongate in concert with polarized basal actin and monopolar protrusions in FCs. An illustration of the basal surface of a stage 8 FC highlights planar polarized features including monopolar actin-rich protrusions and localized Fat2, as compared to Dlar and βPS Integrin.

(A) Live imaging of rotating st. 7 follicle at different time points. Volocity-rendered 3D image; marked cells are pseudocolored. (B) Still image stained as in 1A emphasizing that neighboring follicles can rotate with opposite chiralities. (C) Drawing of basal surface of a rotating follicle, illustrating movement of FCs over stationary ECM fibrils. (D-F) Models for behaviors associated with elongating follicles: tissue rotation organizing a planar polarized collagen corset that channels growth along the A-P axis (D), oscillating contractions providing further force resistance in the follicle center (E), and collagen corset maintaining anisotropic forces that have driven egg elongation (F). Fig. 4D was modified from Haigo and Bilder (2011).

In the abdomen (A), subtle asymmetries in the number of Ft-Ds heterodimers on each side of a cell result in asymmetric localization of Dachs; whether polarity propagation occurs simultaneously or sequentially is unclear. In the wing (B), molecular interactions between PCP signaling components, both reinforcing (Fz and Vang complexes across cell boundaries) and exclusionary (Fz and Vang complexes within a cell) create a feedback loop that amplifies and coordinates polarity. In the germband (C), planar polarization of actin filaments and Myosin II, along with complementary asymmetries of cell-junction proteins, drive polarized junctional remodeling and cell intercalation. In contrast to these stationary epithelia, in motile FCs (D) mechanotransduction between chirally polarized integrin-mediated adhesion complexes and the extracellular matrix amplifies polarity, while rotation of the epithelium coordinates it. Numbers denote a temporal sequence of events; question marks denote ambiguity in temporal sequence. See text for details.
References
-
- Adams DS, Keller R, Koehl MAR. The Mechanics of Notochord Elongation, Straightening and Stiffening in the Embryo of Xenopus-Laevis. Development. 1990;110:115–130. - PubMed
-
- Aigouy B, Farhadifar R, Staple DB, Sagner A, Roper JC, Julicher F, Eaton S. Cell flow reorients the axis of planar polarity in the wing epithelium of Drosophila. Cell. 2010;142:773–786. - PubMed
-
- Axelrod JD. Progress and challenges in understanding planar cell polarity signaling. Semin Cell Dev Biol. 2009;20:964–971. - PubMed
-
- Baskin TI. Anisotropic expansion of the plant cell wall. Annu Rev Cell Dev Biol. 2005;21:203–222. - PubMed
-
- Bateman J, Reddy R, Saito H, Van Vactor D. The receptor tyrosine phosphatase Dlar and integrins organize actin filaments in the Drosophila follicular epithelium. Curr Biol. 2001;11:1317–1327. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
