

Membrane tension maintains cell polarity by confining signals to the leading edge during neutrophil migration
- PMID: 22265410
- PMCID: PMC3308728
- DOI: 10.1016/j.cell.2011.10.050
Membrane tension maintains cell polarity by confining signals to the leading edge during neutrophil migration
Abstract
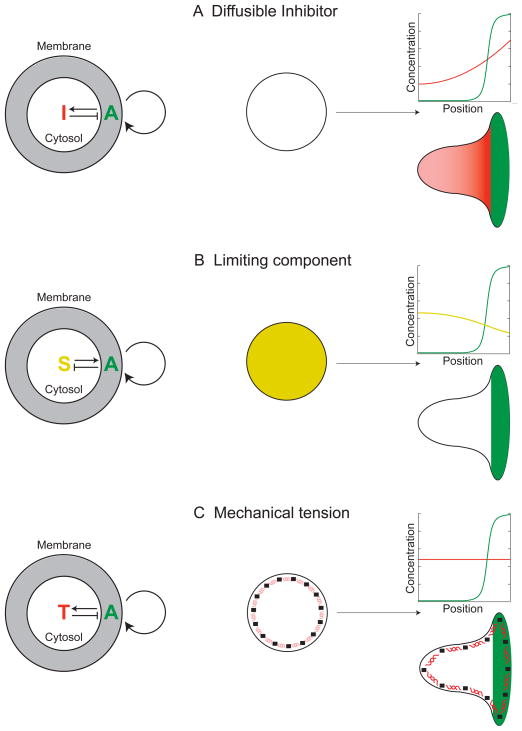
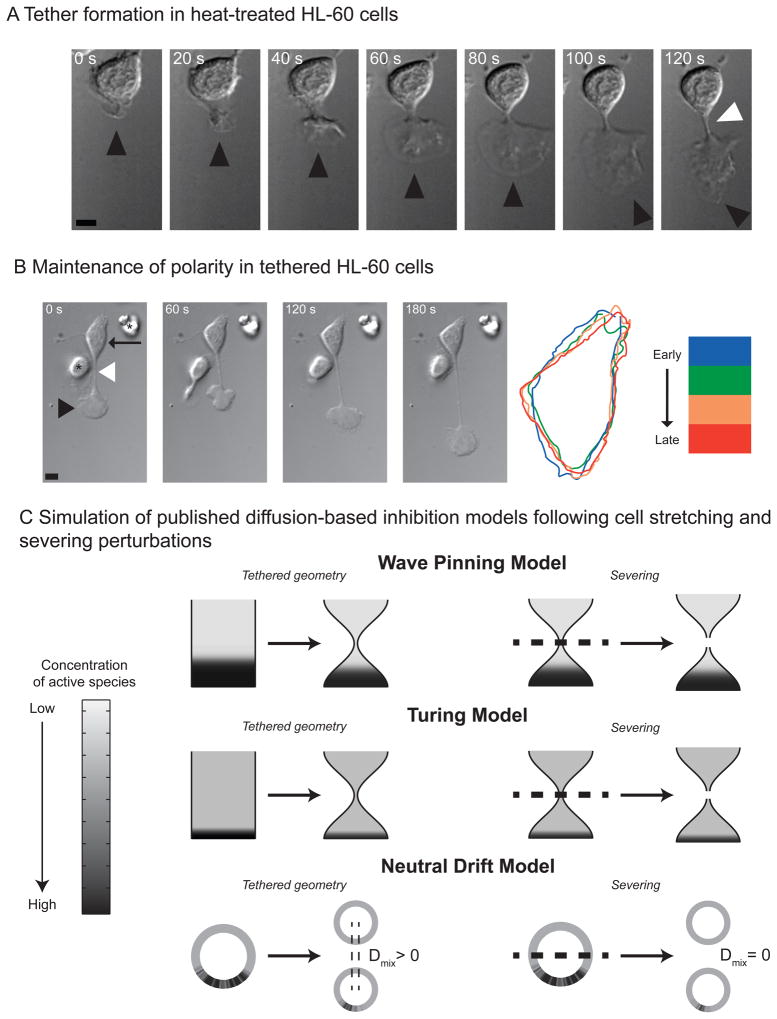
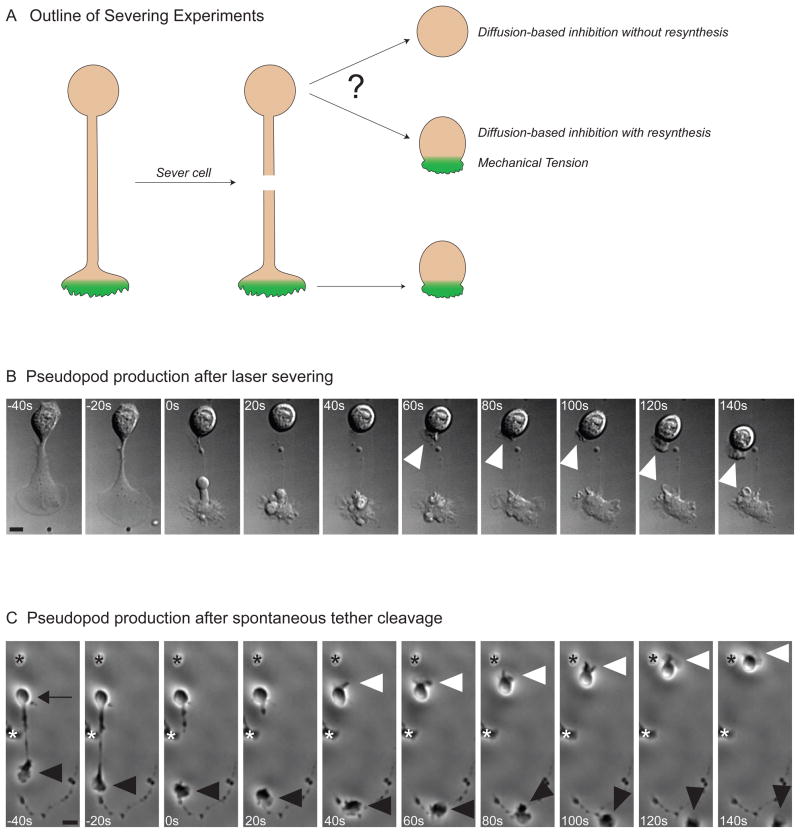
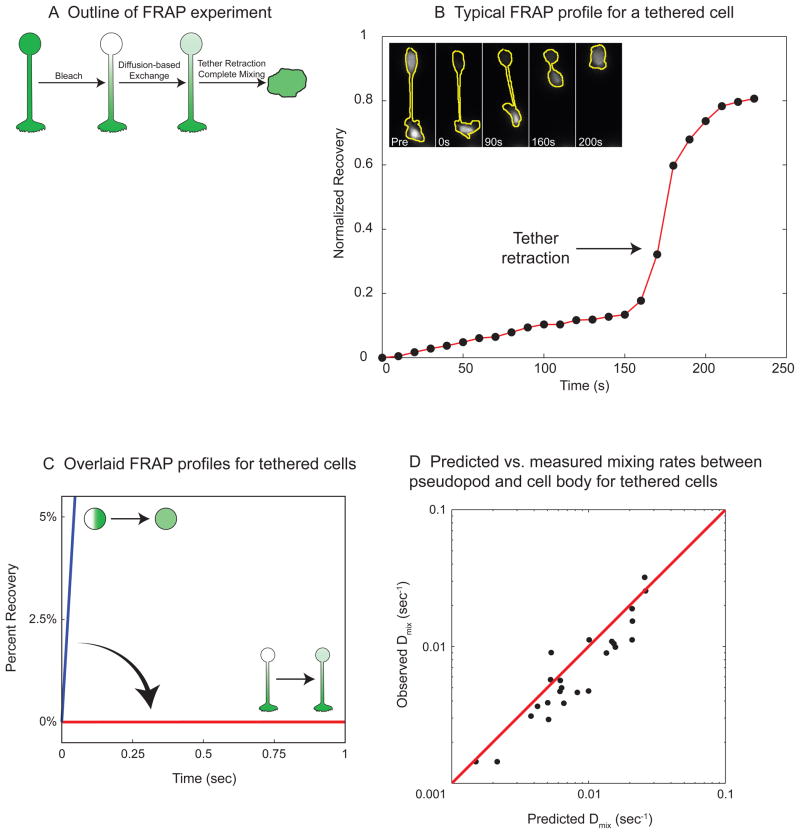
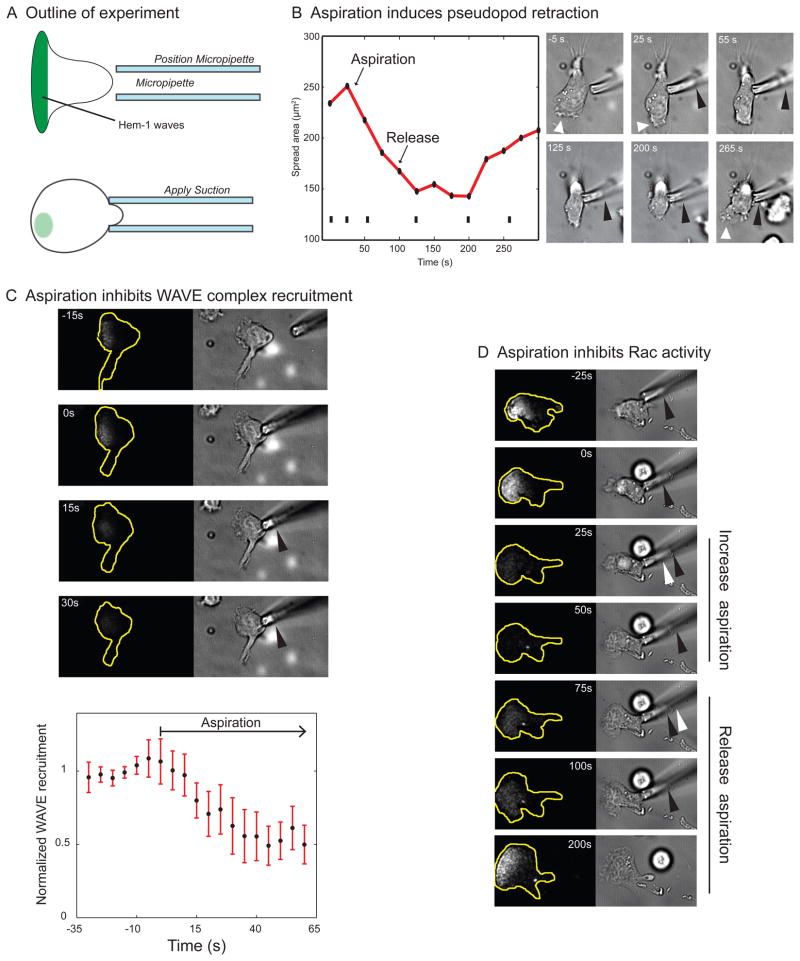
Little is known about how neutrophils and other cells establish a single zone of actin assembly during migration. A widespread assumption is that the leading edge prevents formation of additional fronts by generating long-range diffusible inhibitors or by sequestering essential polarity components. We use morphological perturbations, cell-severing experiments, and computational simulations to show that diffusion-based mechanisms are not sufficient for long-range inhibition by the pseudopod. Instead, plasma membrane tension could serve as a long-range inhibitor in neutrophils. We find that membrane tension doubles during leading-edge protrusion, and increasing tension is sufficient for long-range inhibition of actin assembly and Rac activation. Furthermore, reducing membrane tension causes uniform actin assembly. We suggest that tension, rather than diffusible molecules generated or sequestered at the leading edge, is the dominant source of long-range inhibition that constrains the spread of the existing front and prevents the formation of secondary fronts.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures


Comment in
-
Neutrophils under tension.Dev Cell. 2012 Feb 14;22(2):236-8. doi: 10.1016/j.devcel.2012.01.017. Dev Cell. 2012. PMID: 22340489 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
