

Syk kinase-coupled C-type lectin receptors engage protein kinase C-δ to elicit Card9 adaptor-mediated innate immunity
- PMID: 22265677
- PMCID: PMC3477316
- DOI: 10.1016/j.immuni.2011.11.015
Syk kinase-coupled C-type lectin receptors engage protein kinase C-δ to elicit Card9 adaptor-mediated innate immunity
Abstract
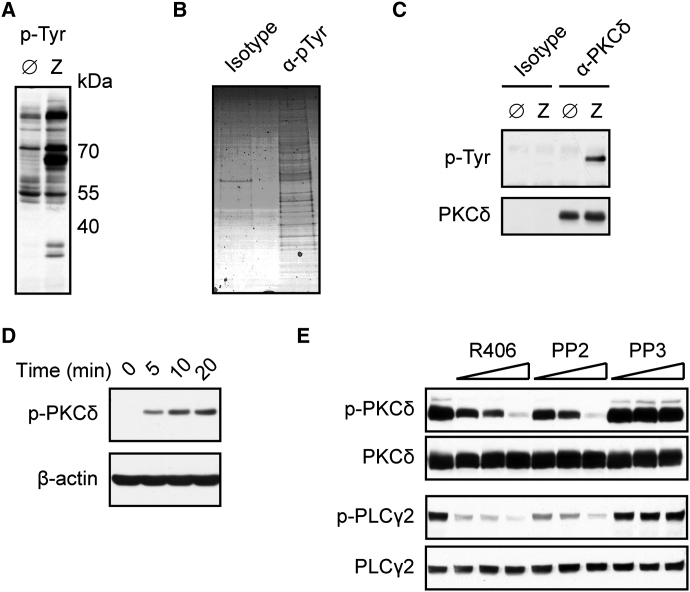
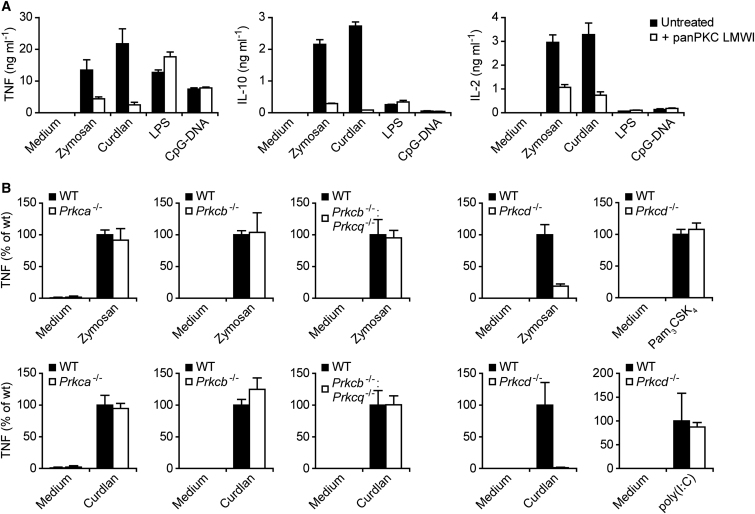
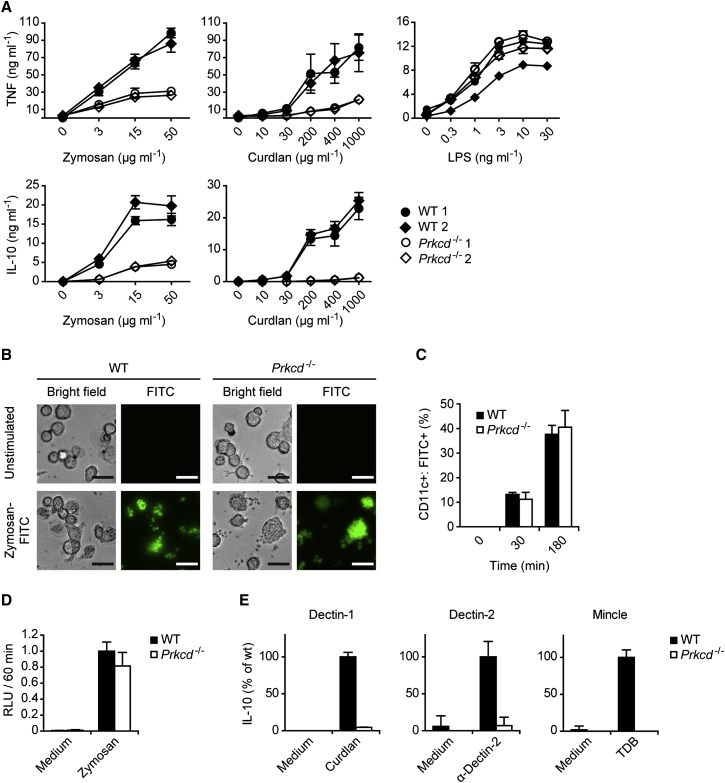
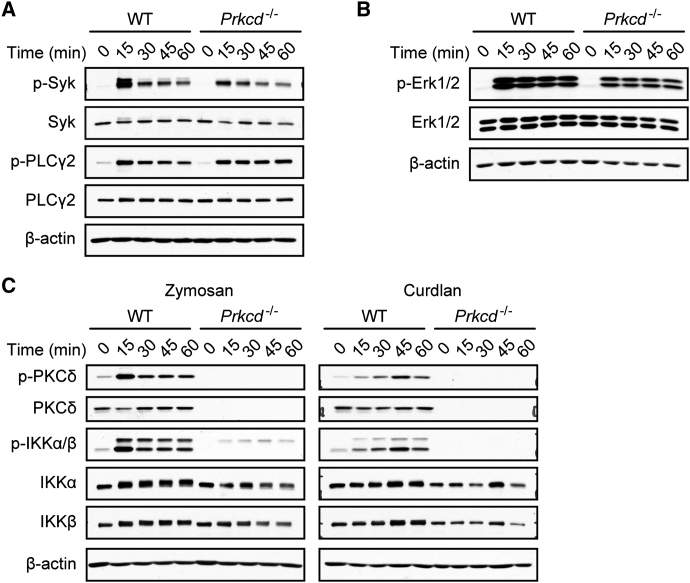
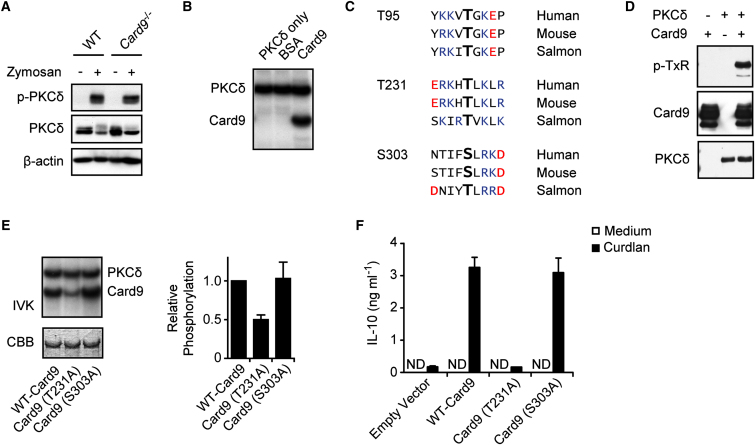
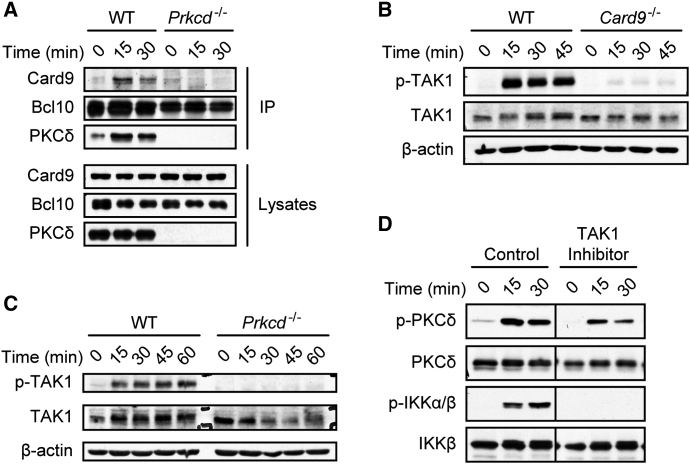
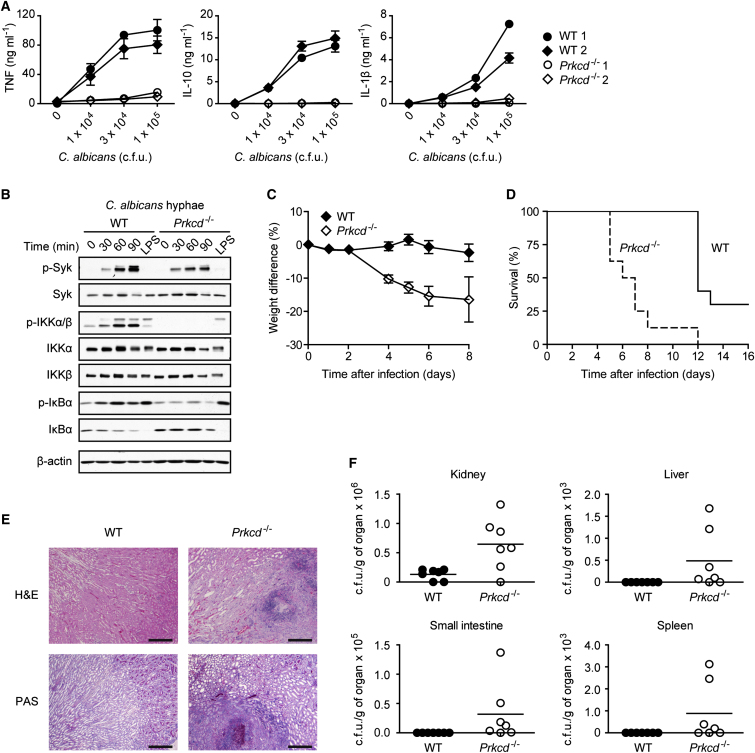
C-type lectin receptors (CLRs) that couple with the kinase Syk are major pattern recognition receptors for the activation of innate immunity and host defense. CLRs recognize fungi and other forms of microbial or sterile danger, and they induce inflammatory responses through the adaptor protein Card9. The mechanisms relaying CLR proximal signals to the core Card9 module are unknown. Here we demonstrated that protein kinase C-δ (PKCδ) was activated upon Dectin-1-Syk signaling, mediated phosphorylation of Card9 at Thr231, and was responsible for Card9-Bcl10 complex assembly and canonical NF-κB control. Prkcd(-/-) dendritic cells, but not those lacking PKCα, PKCβ, or PKCθ, were defective in innate responses to Dectin-1, Dectin-2, or Mincle stimulation. Moreover, Candida albicans-induced cytokine production was blocked in Prkcd(-/-) cells, and Prkcd(-/-) mice were highly susceptible to fungal infection. Thus, PKCδ is an essential link between Syk activation and Card9 signaling for CLR-mediated innate immunity and host protection.
Figures
References
-
- Ariizumi K., Shen G.L., Shikano S., Xu S., Ritter R., 3rd, Kumamoto T., Edelbaum D., Morita A., Bergstresser P.R., Takashima A. Identification of a novel, dendritic cell-associated molecule, dectin-1, by subtractive cDNA cloning. J. Biol. Chem. 2000;275:20157–20167. - PubMed
-
- Blom N., Sicheritz-Pontén T., Gupta R., Gammeltoft S., Brunak S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics. 2004;4:1633–1649. - PubMed
-
- Brown G.D., Gordon S. Immune recognition. A new receptor for beta-glucans. Nature. 2001;413:36–37. - PubMed
-
- Chen S.T., Lin Y.L., Huang M.T., Wu M.F., Cheng S.C., Lei H.Y., Lee C.K., Chiou T.W., Wong C.H., Hsieh S.L. CLEC5A is critical for dengue-virus-induced lethal disease. Nature. 2008;453:672–676. - PubMed
-
- Choudhary C., Olsen J.V., Brandts C., Cox J., Reddy P.N., Böhmer F.D., Gerke V., Schmidt-Arras D.E., Berdel W.E., Müller-Tidow C. Mislocalized activation of oncogenic RTKs switches downstream signaling outcomes. Mol. Cell. 2009;36:326–339. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
