

Kras(G12D) and p53 mutation cause primary intrahepatic cholangiocarcinoma
- PMID: 22266220
- PMCID: PMC3306549
- DOI: 10.1158/0008-5472.CAN-11-3596
Kras(G12D) and p53 mutation cause primary intrahepatic cholangiocarcinoma
Abstract
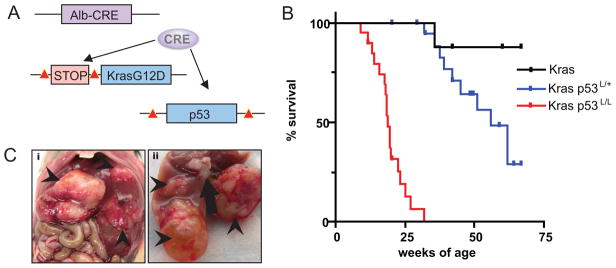
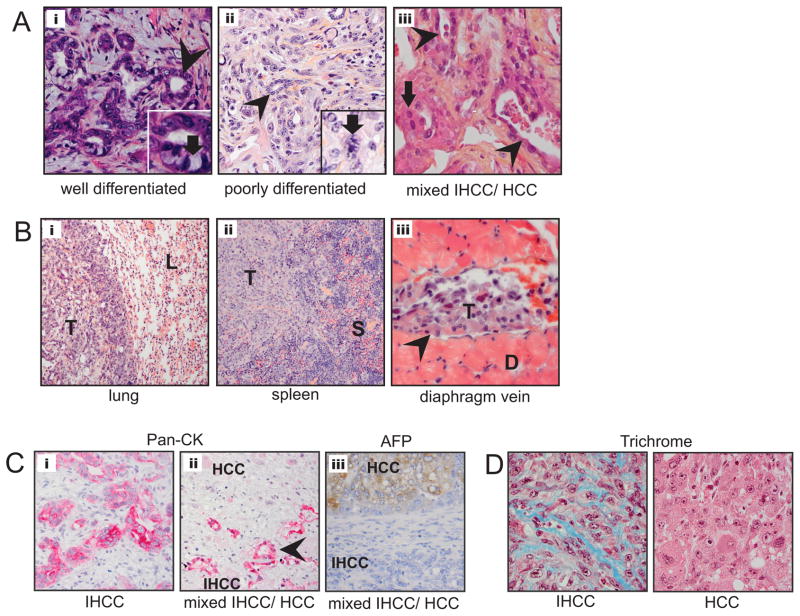
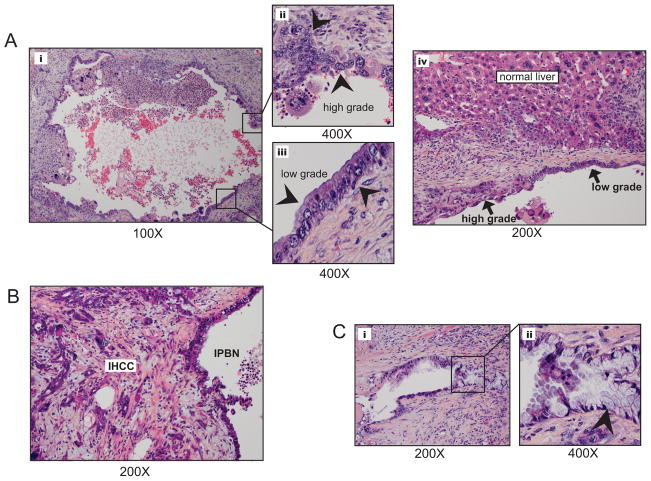
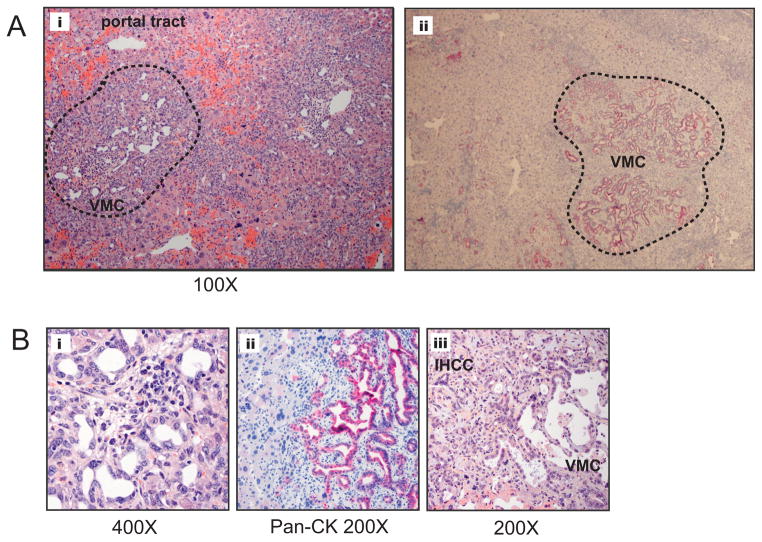
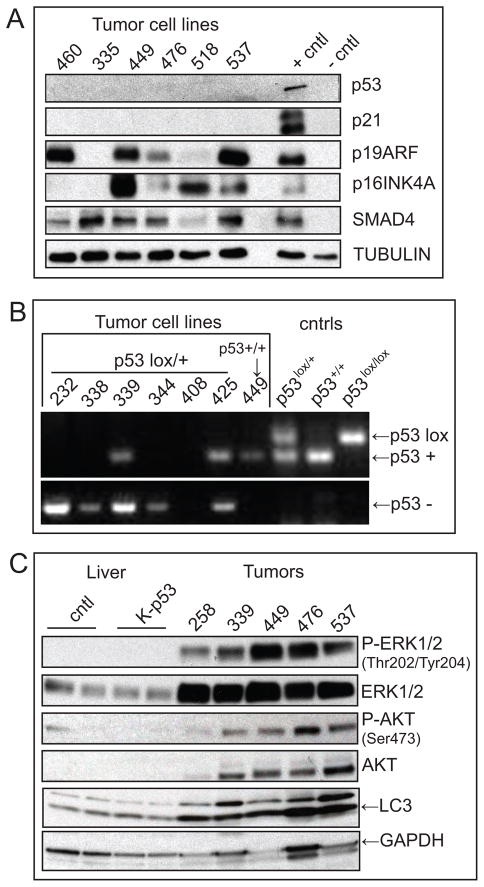
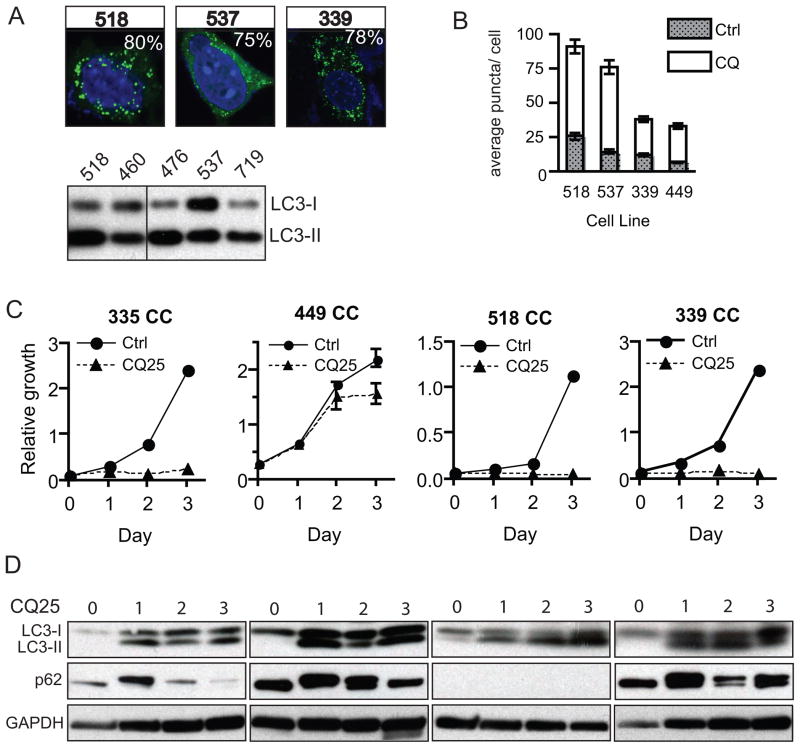
Intrahepatic cholangiocarcinoma (IHCC) is a primary cancer of the liver with an increasing incidence and poor prognosis. Preclinical studies of the etiology and treatment of this disease are hampered by the relatively small number of available IHCC cell lines or genetically faithful animal models. Here we report the development of a genetically engineered mouse model of IHCC that incorporates two of the most common mutations in human IHCC, activating mutations of Kras (Kras(G12D)) and deletion of p53. Tissue-specific activation of Kras(G12D) alone resulted in the development of invasive IHCC with low penetrance and long latency. Latency was shortened by combining Kras(G12D) activation with heterozygous or homozygous deletion of p53 (mean survival of 56 weeks vs. 19 weeks, respectively), which also resulted in widespread local and distant metastasis. Serial analysis showed that the murine models closely recapitulated the multistage histopathologic progression of the human disease, including the development of stroma-rich tumors and the premalignant biliary lesions, intraductal papillary biliary neoplasms (IPBN), and Von Meyenburg complexes (VMC; also known as biliary hamartomas). These findings establish a new genetically and histopathologically faithful model of IHCC and lend experimental support to the hypothesis that IPBN and VMC are precursors to invasive cancers.
Figures
Comment in
-
Autophagy in intra-hepatic cholangiocarcinoma.Autophagy. 2012 Jul 1;8(7):1148-9. doi: 10.4161/auto.20647. Epub 2012 Jul 1. Autophagy. 2012. PMID: 22751196 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
