

Myeloid translocation gene 16 is required for maintenance of haematopoietic stem cell quiescence
- PMID: 22266796
- PMCID: PMC3321173
- DOI: 10.1038/emboj.2011.500
Myeloid translocation gene 16 is required for maintenance of haematopoietic stem cell quiescence
Abstract
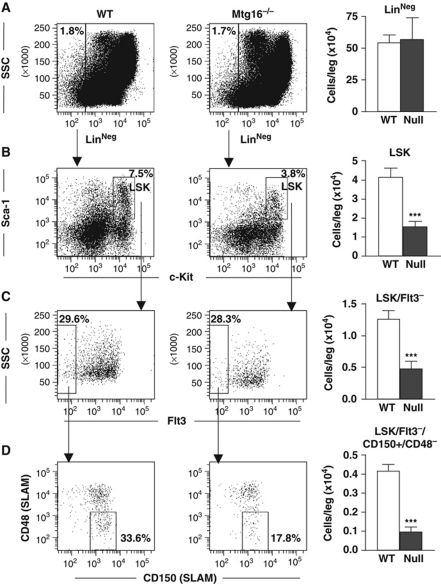
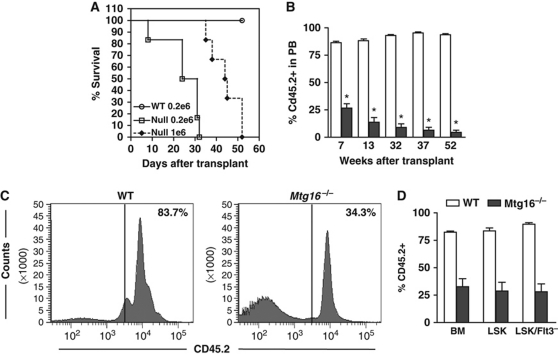
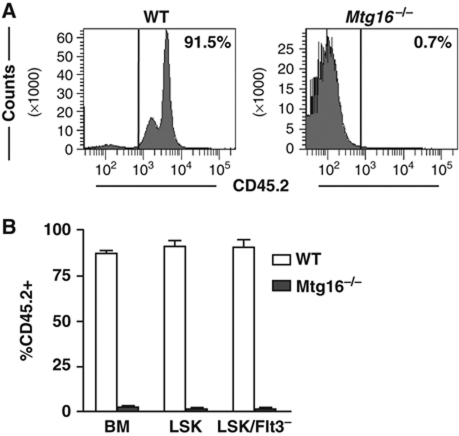
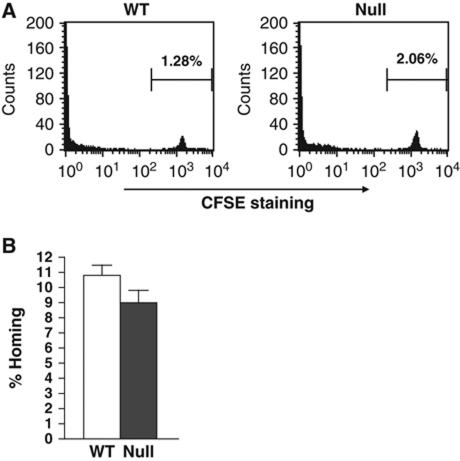
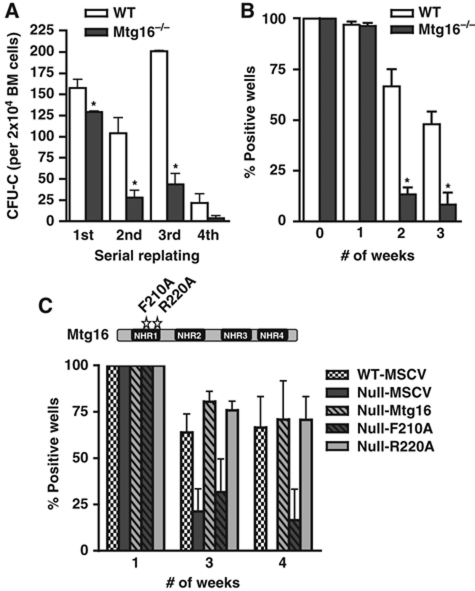
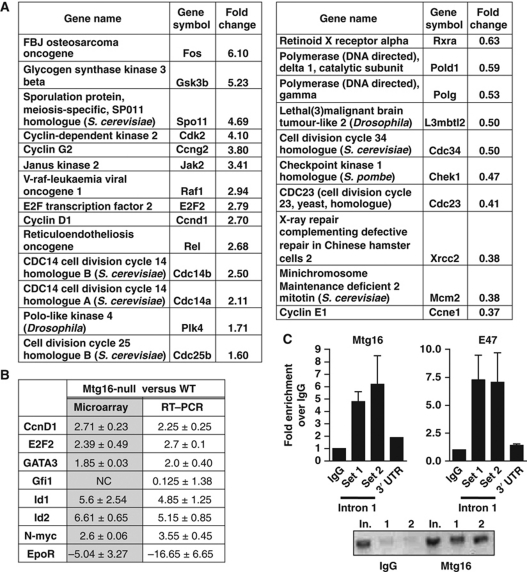
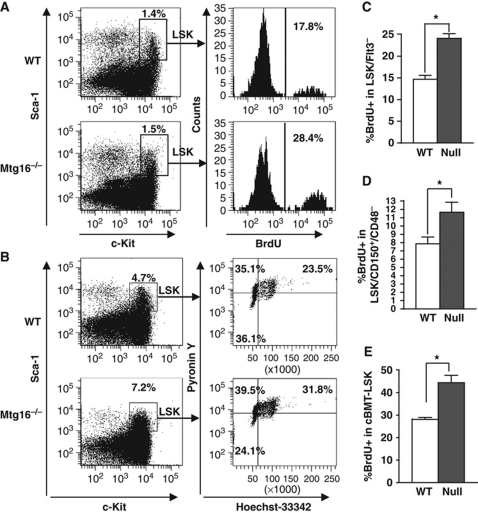
The t(8;21) and t(16;21) that are associated with acute myeloid leukaemia disrupt two closely related genes termed Myeloid Translocation Genes 8 (MTG8) and 16 (MTG16), respectively. Many of the transcription factors that recruit Mtg16 regulate haematopoietic stem and progenitor cell functions and are required to maintain stem cell self-renewal potential. Accordingly, we found that Mtg16-null bone marrow (BM) failed in BM transplant assays. Moreover, when removed from the animal, Mtg16-deficient stem cells continued to show defects in stem cell self-renewal assays, suggesting a requirement for Mtg16 in this process. Gene expression analysis indicated that Mtg16 was required to suppress the expression of several key cell-cycle regulators including E2F2, and chromatin immunoprecipitation assays detected Mtg16 near an E2A binding site within the first intron of E2F2. BrdU incorporation assays indicated that in the absence of Mtg16 more long-term stem cells were in the S phase, even after competitive BM transplantation where normal stem and progenitor cells are present, suggesting that Mtg16 plays a role in the maintenance of stem cell quiescence.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Amann JM, Chyla BJ, Ellis TC, Martinez A, Moore AC, Franklin JL, McGhee L, Meyers S, Ohm JE, Luce KS, Ouelette AJ, Washington MK, Thompson MA, King D, Gautam S, Coffey RJ, Whitehead RH, Hiebert SW (2005) Mtgr1 is a transcriptional corepressor that is required for maintenance of the secretory cell lineage in the small intestine. Mol Cell Biol 25: 9576–9585 - PMC - PubMed
-
- Boggs DR (1984) The total marrow mass of the mouse: a simplified method of measurement. Am J Hematol 16: 277–286 - PubMed
-
- Buitenhuis M, van Deutekom HW, Verhagen LP, Castor A, Jacobsen SE, Lammers JW, Koenderman L, Coffer PJ (2005) Differential regulation of granulopoiesis by the basic helix-loop-helix transcriptional inhibitors Id1 and Id2. Blood 105: 4272–4281 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P30CA68485/CA/NCI NIH HHS/United States
- 5P30DK58404/DK/NIDDK NIH HHS/United States
- R01-HL088494/HL/NHLBI NIH HHS/United States
- P30 DK058404/DK/NIDDK NIH HHS/United States
- P30 CA068485/CA/NCI NIH HHS/United States
- F30 HL093993/HL/NHLBI NIH HHS/United States
- UL1 RR024975/RR/NCRR NIH HHS/United States
- R01-CA64140/CA/NCI NIH HHS/United States
- UL1 RR024975-01/RR/NCRR NIH HHS/United States
- R01 CA112005/CA/NCI NIH HHS/United States
- R01-CA112005/CA/NCI NIH HHS/United States
- R01 HL088494/HL/NHLBI NIH HHS/United States
- R01 CA064140/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
