

TOR complex 2 (TORC2) in Dictyostelium suppresses phagocytic nutrient capture independently of TORC1-mediated nutrient sensing
- PMID: 22266904
- PMCID: PMC3269021
- DOI: 10.1242/jcs.077040
TOR complex 2 (TORC2) in Dictyostelium suppresses phagocytic nutrient capture independently of TORC1-mediated nutrient sensing
Abstract
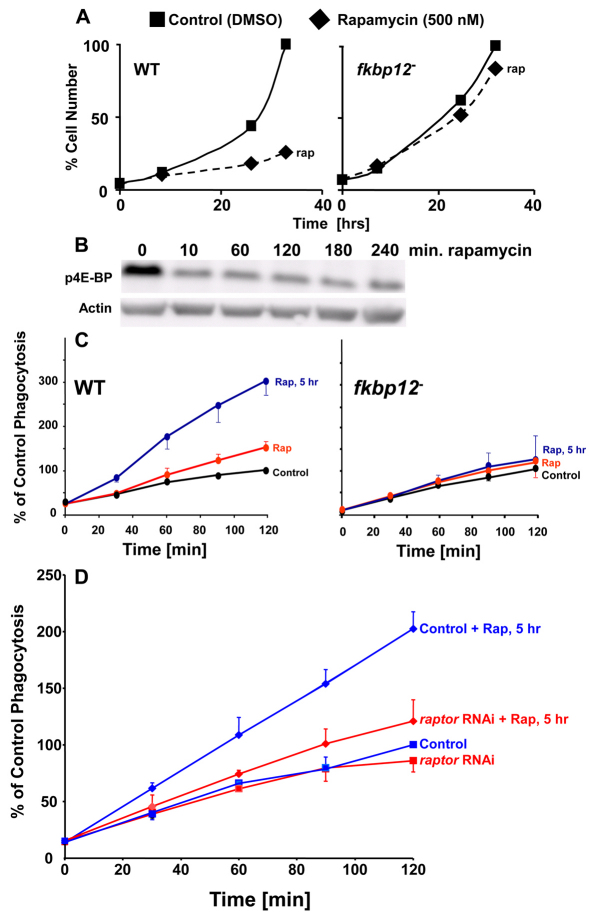
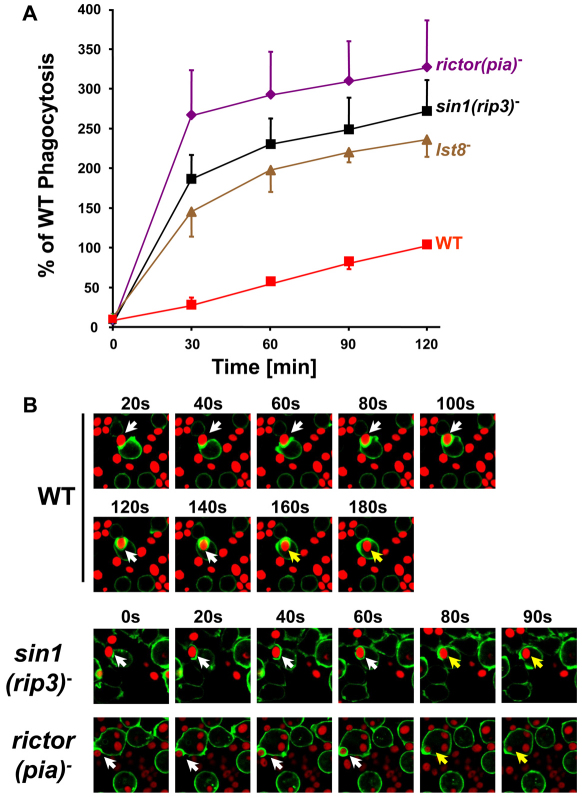
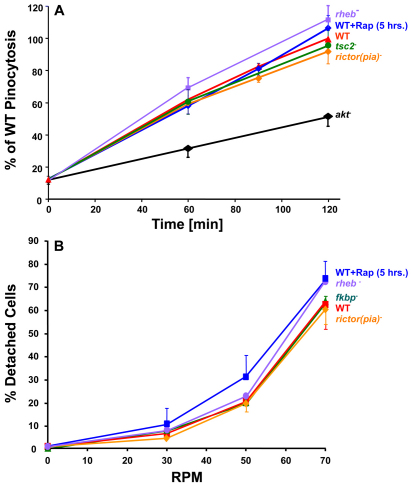
The TOR protein kinase functions in two distinct complexes, TOR complex 1 (TORC1) and 2 (TORC2). TORC1 is required for growth in response to growth factors, nutrients and the cellular energy state; TORC2 regulates AKT signaling, which can modulate cytoskeletal polarization. In its ecological niche, Dictyostelium engulf bacteria and yeast for nutrient capture. Despite the essential role of TORC1 in control of cellular growth, we show that nutrient particle capture (phagocytosis) in Dictyostelium is independent of TORC1-mediated nutrient sensing and growth regulation. However, loss of Dictyostelium TORC2 components Rictor/Pia, SIN1/RIP3 and Lst8 promotes nutrient particle uptake; inactivation of TORC2 leads to increased efficiency and speed of phagocytosis. In contrast to phagocytosis, we show that macropinocytosis, an AKT-dependent process for cellular uptake of fluid phase nutrients, is not regulated by either of the TOR complexes. The integrated and balanced regulation of TORC1 and TORC2 might be crucial in Dictyostelium to coordinate growth and energy needs with other essential TOR-regulated processes.
Figures





Similar articles
-
Target of Rapamycin (TOR) Regulates Growth in Response to Nutritional Signals.Microbiol Spectr. 2016 Oct;4(5). doi: 10.1128/microbiolspec.FUNK-0006-2016. Microbiol Spectr. 2016. PMID: 27763256 Review.
-
Isp7 is a novel regulator of amino acid uptake in the TOR signaling pathway.Mol Cell Biol. 2014 Mar;34(5):794-806. doi: 10.1128/MCB.01473-13. Epub 2013 Dec 16. Mol Cell Biol. 2014. PMID: 24344203 Free PMC article.
-
Amino acid signaling in TOR activation.Annu Rev Biochem. 2011;80:1001-32. doi: 10.1146/annurev-biochem-062209-094414. Annu Rev Biochem. 2011. PMID: 21548787 Review.
-
LST8 regulates cell growth via target-of-rapamycin complex 2 (TORC2).Mol Cell Biol. 2012 Jun;32(12):2203-13. doi: 10.1128/MCB.06474-11. Epub 2012 Apr 9. Mol Cell Biol. 2012. PMID: 22493059 Free PMC article.
-
Identification of Sin1 as an essential TORC2 component required for complex formation and kinase activity.Genes Dev. 2006 Oct 15;20(20):2820-32. doi: 10.1101/gad.1461206. Genes Dev. 2006. PMID: 17043309 Free PMC article.
Cited by
-
Phosphatidic acid and lipid-sensing by mTOR.Trends Endocrinol Metab. 2013 Jun;24(6):272-8. doi: 10.1016/j.tem.2013.02.003. Epub 2013 Mar 16. Trends Endocrinol Metab. 2013. PMID: 23507202 Free PMC article.
-
CRISPR/Cas9-based genome-wide screening of Dictyostelium.Sci Rep. 2022 Jul 2;12(1):11215. doi: 10.1038/s41598-022-15500-3. Sci Rep. 2022. PMID: 35780186 Free PMC article.
-
Enhanced Sestrin expression through Tanshinone 2A treatment improves PI3K-dependent inhibition of glioma growth.Cell Death Discov. 2023 May 19;9(1):172. doi: 10.1038/s41420-023-01462-6. Cell Death Discov. 2023. PMID: 37202382 Free PMC article.
-
Methods to Monitor and Quantify Autophagy in the Social Amoeba Dictyostelium discoideum.Cells. 2017 Jul 3;6(3):18. doi: 10.3390/cells6030018. Cells. 2017. PMID: 28671610 Free PMC article. Review.
-
Ras, PI3K and mTORC2 - three's a crowd?J Cell Sci. 2020 Oct 8;133(19):jcs234930. doi: 10.1242/jcs.234930. J Cell Sci. 2020. PMID: 33033115 Free PMC article. Review.
References
-
- Abraham R. T., Wiederrecht G. J. (1996). Immunopharmacology of rapamycin. Annu. Rev. Immunol. 14, 483-510 - PubMed
-
- Aspuria P. J., Sato T., Tamanoi F. (2007). The TSC/Rheb/TOR signaling pathway in fission yeast and mammalian cells: temperature sensitive and constitutive active mutants of TOR. Cell Cycle 6, 1692-1695 - PubMed
-
- Blaauw M., Linskens M. H., van Haastert P. J. (2000). Efficient control of gene expression by a tetracycline-dependent transactivator in single Dictyostelium discoideum cells. Gene 252, 71-82 - PubMed
-
- Brunn G. J., Hudson C. C., Sekulic A., Williams J. M., Hosoi H., Houghton P. J., Lawrence J. C., Jr, Abraham R. T. (1997). Phosphorylation of the translational repressor PHAS-I by the mammalian target of rapamycin. Science 277, 99-101 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
