

Periostin modulates myofibroblast differentiation during full-thickness cutaneous wound repair
- PMID: 22266908
- PMCID: PMC3269025
- DOI: 10.1242/jcs.087841
Periostin modulates myofibroblast differentiation during full-thickness cutaneous wound repair
Abstract
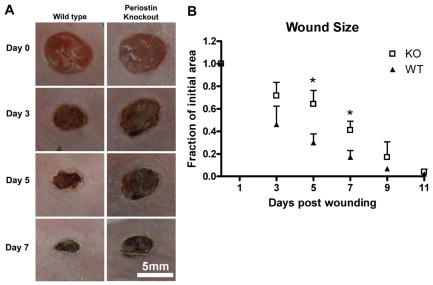
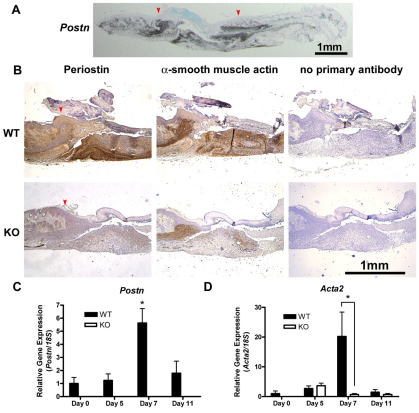
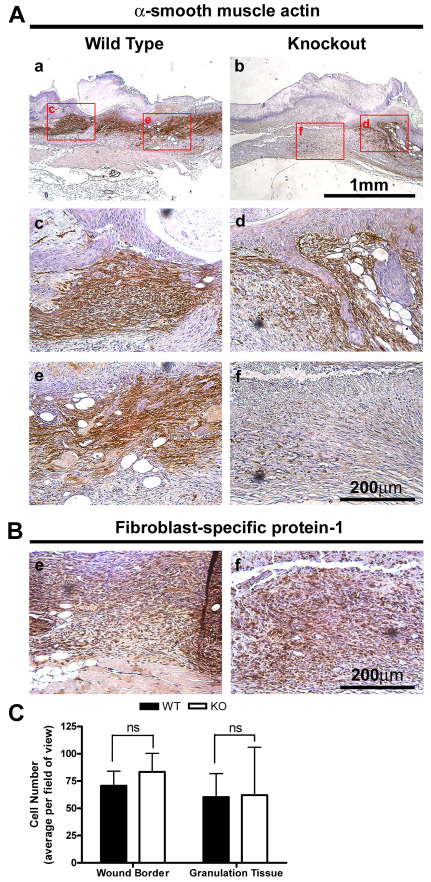
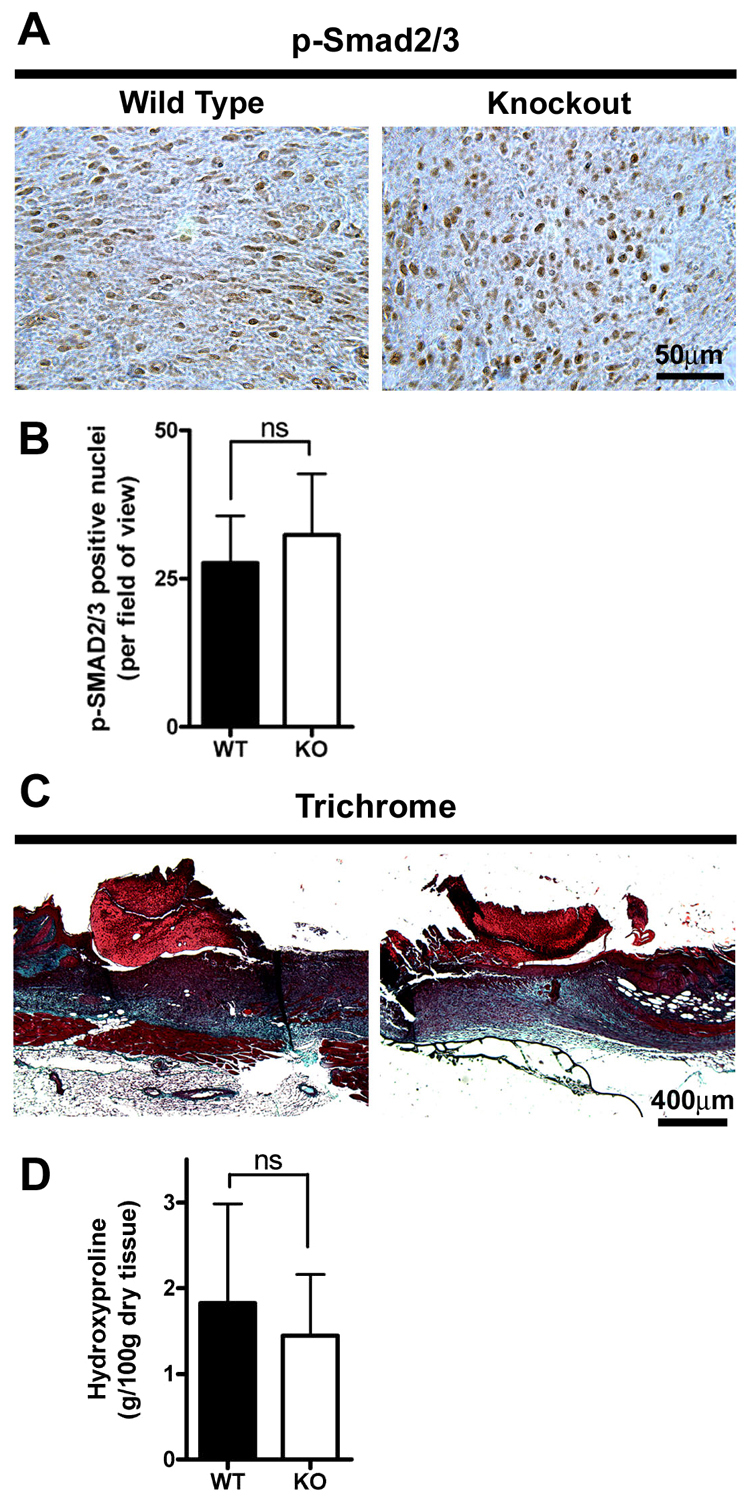
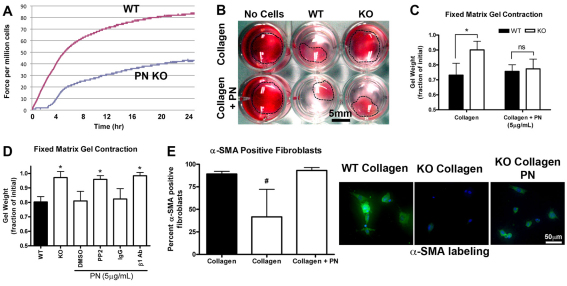
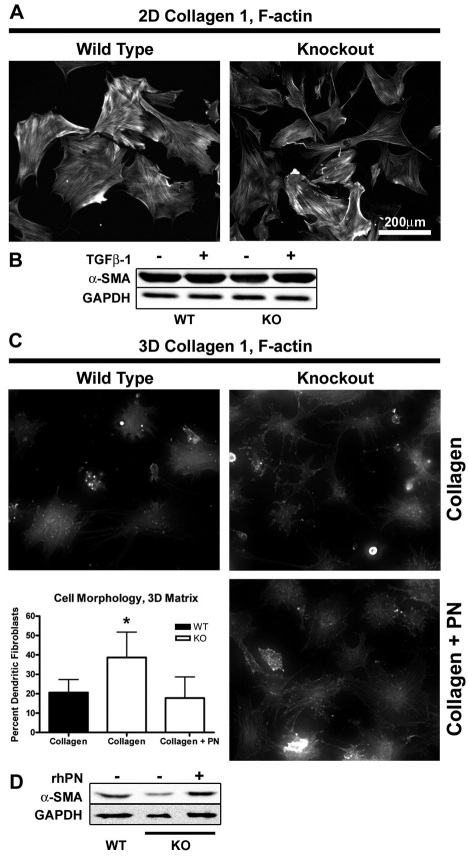
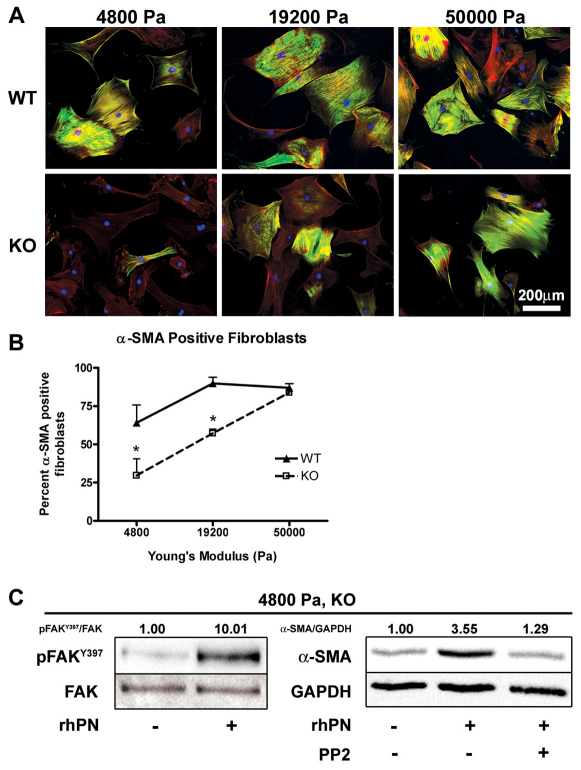
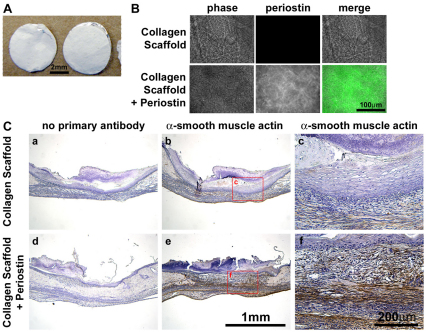
The matricellular protein periostin is expressed in the skin. Although periostin has been hypothesized to contribute to dermal homeostasis and repair, this has not been directly tested. To assess the contribution of periostin to dermal healing, 6 mm full-thickness excisional wounds were created in the skin of periostin-knockout and wild-type, sex-matched control mice. In wild-type mice, periostin was potently induced 5-7 days after wounding. In the absence of periostin, day 7 wounds showed a significant reduction in myofibroblasts, as visualized by expression of α-smooth muscle actin (α-SMA) within the granulation tissue. Delivery of recombinant human periostin by electrospun collagen scaffolds restored α-SMA expression. Isolated wild-type and knockout dermal fibroblasts did not differ in in vitro assays of adhesion or migration; however, in 3D culture, periostin-knockout fibroblasts showed a significantly reduced ability to contract a collagen matrix, and adopted a dendritic phenotype. Recombinant periostin restored the defects in cell morphology and matrix contraction displayed by periostin-deficient fibroblasts in a manner that was sensitive to a neutralizing anti-β1-integrin and to the FAK and Src inhibitor PP2. We propose that periostin promotes wound contraction by facilitating myofibroblast differentiation and contraction.
Figures
References
-
- Aplin J. D., Hughes R. C. (1981). Protein-derivatised glass coverslips for the study of cell-to substratum adhesion. Anal. Biochem. 113, 144-148 - PubMed
-
- Babu M., Diegelmann R., Oliver N. (1992). Keloid fibroblasts exhibit an altered response to TGF-beta. J. Invest. Dermatol. 99, 650-655 - PubMed
-
- Bao S., Ouyang G., Bai X., Huang Z., Ma C., Liu M., Shao R., Anderson R. M., Rich J. N., Wang X. F. (2004). Periostin potently promotes metastatic growth of colon cancer by augmenting cell survival via the Akt/PKB pathway. Cancer Cell 5, 329-339 - PubMed
-
- Baril P., Gangeswaran R., Mahon P. C., Caulee K., Kocher H. M., Harada T., Zhu M., Kalthoff H., Crnogorac-Jurcevic T., Lemoine N. R. (2007). Periostin promotes invasiveness and resistance of pancreatic cancer cells to hypoxia-induced cell death: role of the beta4 integrin and the PI3k pathway. Oncogene 26, 2082-2094 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
