

Structure of complement C6 suggests a mechanism for initiation and unidirectional, sequential assembly of membrane attack complex (MAC)
- PMID: 22267737
- PMCID: PMC3323040
- DOI: 10.1074/jbc.M111.327809
Structure of complement C6 suggests a mechanism for initiation and unidirectional, sequential assembly of membrane attack complex (MAC)
Abstract
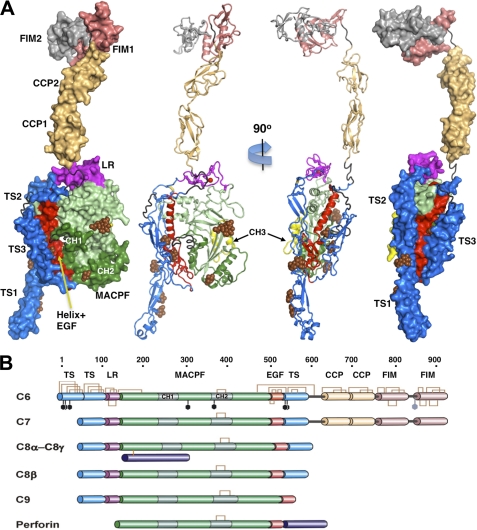
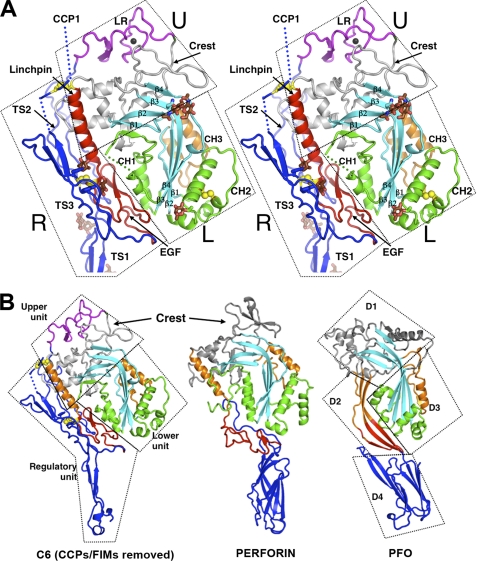
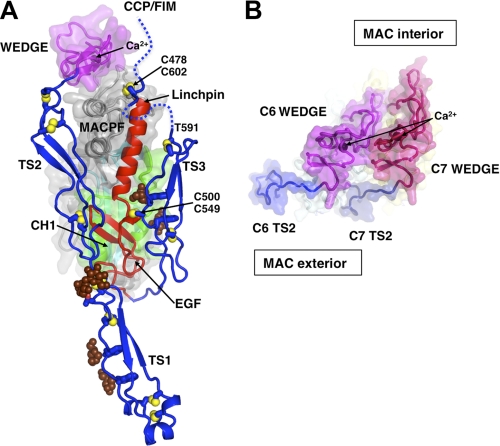
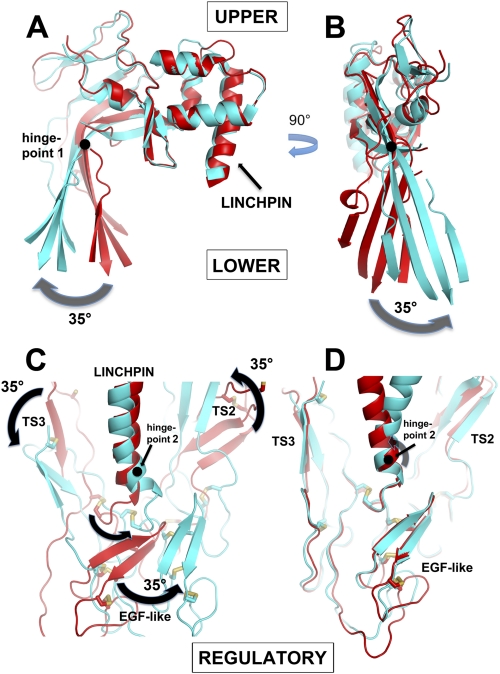
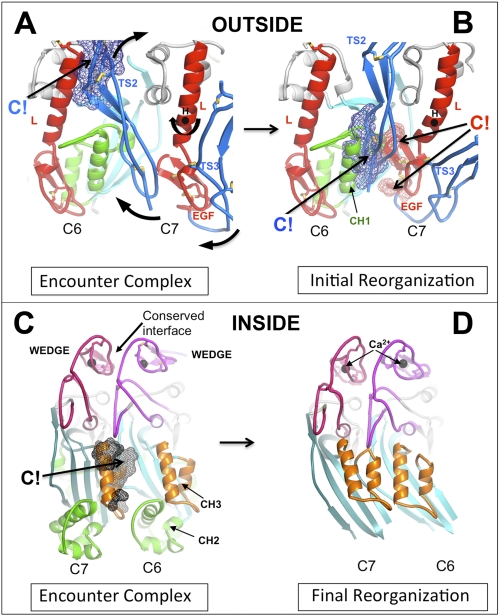
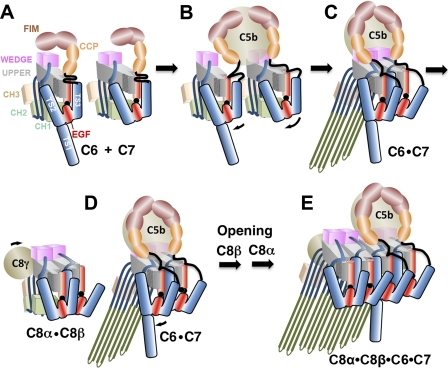
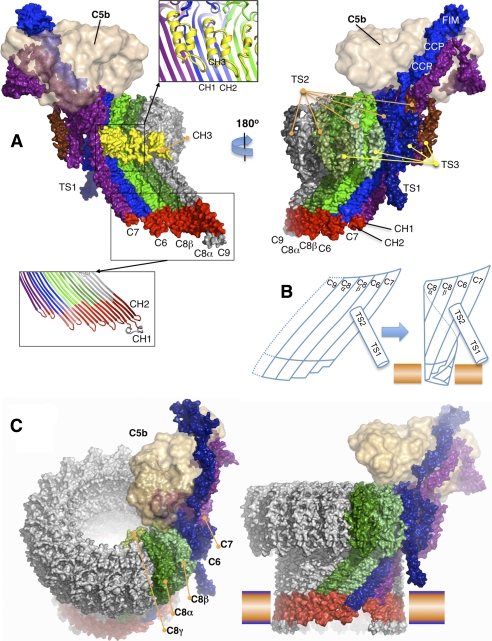
The complement membrane attack complex (MAC) is formed by the sequential assembly of C5b with four homologous proteins as follows: one copy each of C6, C7, and C8 and 12-14 copies of C9. Together these form a lytic pore in bacterial membranes. C6 through C9 comprise a MAC-perforin domain flanked by 4-9 "auxiliary" domains. Here, we report the crystal structure of C6, the first and longest of the pore proteins to be recruited by C5b. Comparisons with the structures of the C8αβγ heterodimer and perforin show that the central domain of C6 adopts a "closed" (perforin-like) state that is distinct from the "open" conformations in C8. We further show that C6, C8α, and C8β contain three homologous subdomains ("upper," "lower," and "regulatory") related by rotations about two hinge points. In C6, the regulatory segment includes four auxiliary domains that stabilize the closed conformation, inhibiting release of membrane-inserting elements. In C8β, rotation of the regulatory segment is linked to an opening of the central β-sheet of its clockwise partner, C8α. Based on these observations, we propose a model for initiation and unidirectional propagation of the MAC in which the auxiliary domains play key roles: in the assembly of the C5b-8 initiation complex; in driving and regulating the opening of the β-sheet of the MAC-performin domain of each new recruit as it adds to the growing pore; and in stabilizing the final pore. Our model of the assembled pore resembles those of the cholesterol-dependent cytolysins but is distinct from that recently proposed for perforin.
Figures
References
-
- Kimura A., Ikeo K., Nonaka M. (2009) Evolutionary origin of the vertebrate blood complement and coagulation systems inferred from liver EST analysis of lamprey. Dev. Comp. Immunol. 33, 77–87 - PubMed
-
- DiScipio R. G. (1998) in The Complement System (Rother K., Till G. O., Hansch G. M., eds) 2 Ed., pp. 50–68, Springer-Verlag, New York
-
- Müller-Eberhard H. J. (1986) The membrane attack complex of complement. Annu. Rev. Immunol. 4, 503–528 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
