

Biochemical characterization of ribosome assembly GTPase RbgA in Bacillus subtilis
- PMID: 22267738
- PMCID: PMC3318688
- DOI: 10.1074/jbc.M111.331322
Biochemical characterization of ribosome assembly GTPase RbgA in Bacillus subtilis
Abstract
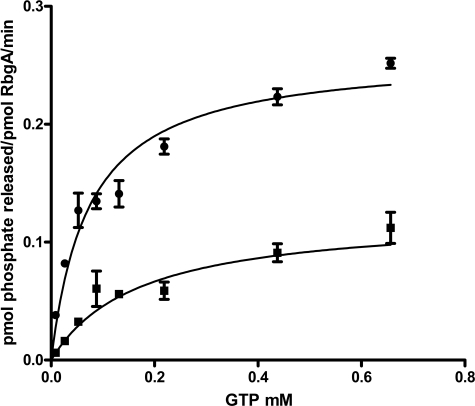
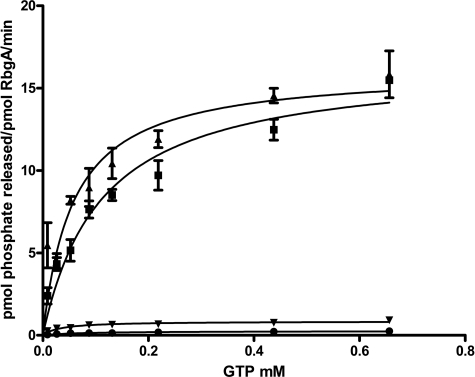
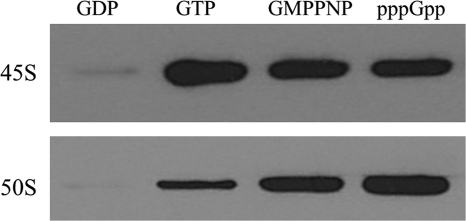
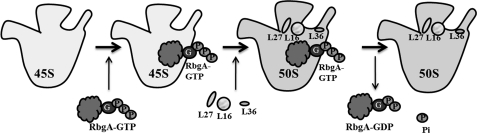
The ribosome biogenesis GTPase A protein RbgA is involved in the assembly of the large ribosomal subunit in Bacillus subtilis, and homologs of RbgA are implicated in the biogenesis of mitochondrial, chloroplast, and cytoplasmic ribosomes in archaea and eukaryotes. The precise function of how RbgA contributes to ribosome assembly is not understood. Defects in RbgA give rise to a large ribosomal subunit that is immature and migrates at 45 S in sucrose density gradients. Here, we report a detailed biochemical analysis of RbgA and its interaction with the ribosome. We found that RbgA, like most other GTPases, exhibits a very slow k(cat) (14 h(-1)) and has a high K(m) (90 μM). Homology modeling of the RbgA switch I region using the K-loop GTPase MnmE as a template suggested that RbgA requires K(+) ions for GTPase activity, which was confirmed experimentally. Interaction with 50 S subunits, but not 45 S intermediates, increased GTPase activity by ∼55-fold. Stable association with 50 S subunits and 45 S intermediates was nucleotide-dependent, and GDP did not support strong interaction with either of the subunits. GTP and guanosine 5'-(β,γ-imido)triphosphate (GMPPNP) were sufficient to promote association with the 45 S intermediate, whereas only GMPPNP was able to support binding to the 50 S subunit, presumably due to the stimulation of GTP hydrolysis. These results support a model in which RbgA promotes a late step in ribosome biogenesis and that one role of GTP hydrolysis is to stimulate dissociation of RbgA from the ribosome.
Figures

References
-
- Nierhaus K. H. (1991) The assembly of prokaryotic ribosomes. Biochimie 73, 739–755 - PubMed
-
- Nomura M., Erdmann V. A. (1970) Reconstitution of 50 S ribosomal subunits from dissociated molecular components. Nature 228, 744–748 - PubMed
-
- Dez C., Tollervey D. (2004) Ribosome synthesis meets the cell cycle. Curr. Opin. Microbiol. 7, 631–637 - PubMed
-
- Culver G. M. (2003) Assembly of the 30 S ribosomal subunit. Biopolymers 68, 234–249 - PubMed
-
- Wilson D. N., Nierhaus K. H. (2007) The weird and wonderful world of bacterial ribosome regulation. Crit. Rev. Biochem. Mol. Biol. 42, 187–219 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
