

Structural determinants of Clostridium difficile toxin A glucosyltransferase activity
- PMID: 22267739
- PMCID: PMC3318759
- DOI: 10.1074/jbc.M111.298414
Structural determinants of Clostridium difficile toxin A glucosyltransferase activity
Abstract
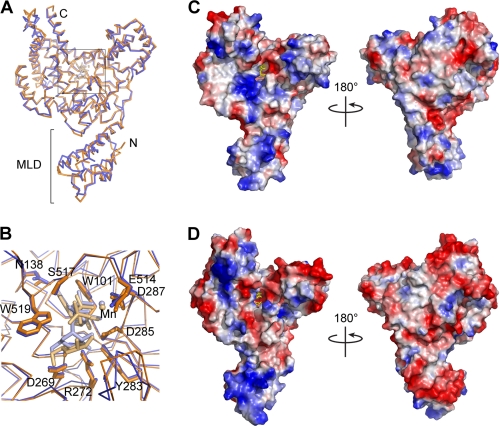
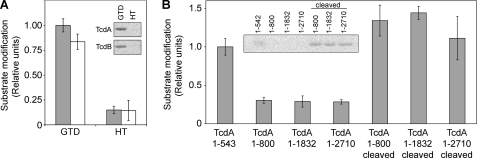
The principle virulence factors in Clostridium difficile pathogenesis are TcdA and TcdB, homologous glucosyltransferases capable of inactivating small GTPases within the host cell. We present crystal structures of the TcdA glucosyltransferase domain in the presence and absence of the co-substrate UDP-glucose. Although the enzymatic core is similar to that of TcdB, the proposed GTPase-binding surface differs significantly. We show that TcdA is comparable with TcdB in its modification of Rho family substrates and that, unlike TcdB, TcdA is also capable of modifying Rap family GTPases both in vitro and in cells. The glucosyltransferase activities of both toxins are reduced in the context of the holotoxin but can be restored with autoproteolytic activation and glucosyltransferase domain release. These studies highlight the importance of cellular activation in determining the array of substrates available to the toxins once delivered into the cell.
Figures


References
-
- Kuehne S. A., Cartman S. T., Heap J. T., Kelly M. L., Cockayne A., Minton N. P. (2010) The role of toxin A and toxin B in Clostridium difficile infection. Nature 467, 711–713 - PubMed
-
- Hofmann F., Busch C., Prepens U., Just I., Aktories K. (1997) Localization of the glucosyltransferase activity of Clostridium difficile toxin B to the N-terminal part of the holotoxin. J. Biol. Chem. 272, 11074–11078 - PubMed
-
- Rupnik M., Pabst S., Rupnik M., von Eichel-Streiber C., Urlaub H., Söling H. D. (2005) Characterization of the cleavage site and function of resulting cleavage fragments after limited proteolysis of Clostridium difficile toxin B (TcdB) by host cells. Microbiology 151, 199–208 - PubMed
-
- von Eichel-Streiber C., Sauerborn M. (1990) Clostridium difficile toxin A carries a C-terminal repetitive structure homologous to the carbohydrate binding region of streptococcal glycosyltransferases. Gene 96, 107–113 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
