

Bile acids via FXR initiate the expression of major transporters involved in the enterohepatic circulation of bile acids in newborn mice
- PMID: 22268101
- PMCID: PMC3362079
- DOI: 10.1152/ajpgi.00370.2011
Bile acids via FXR initiate the expression of major transporters involved in the enterohepatic circulation of bile acids in newborn mice
Abstract
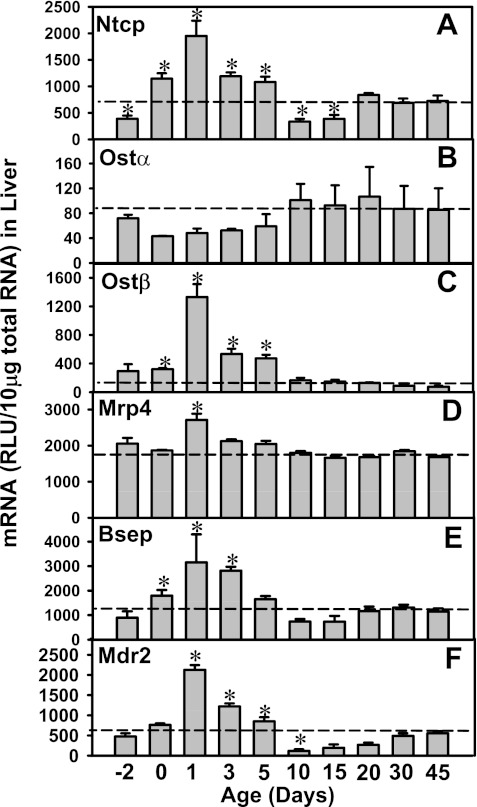
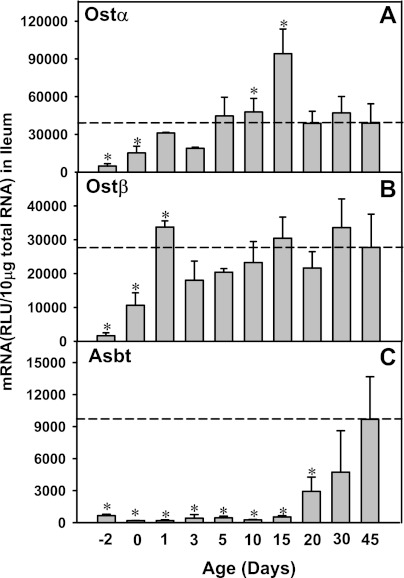
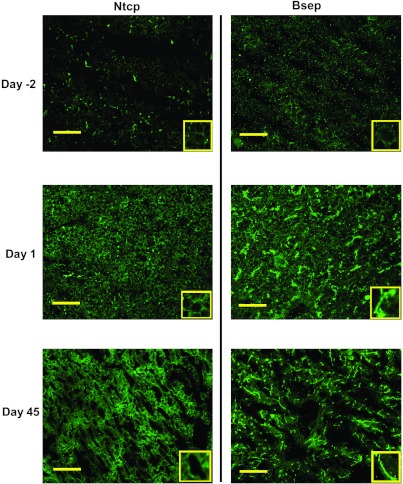
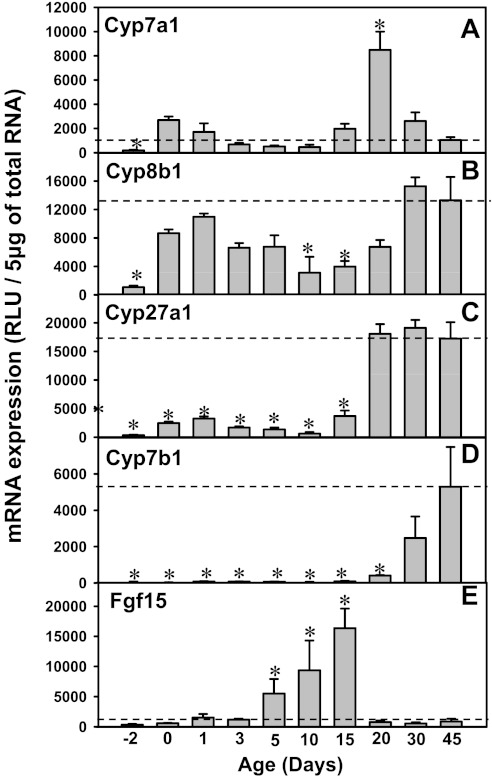
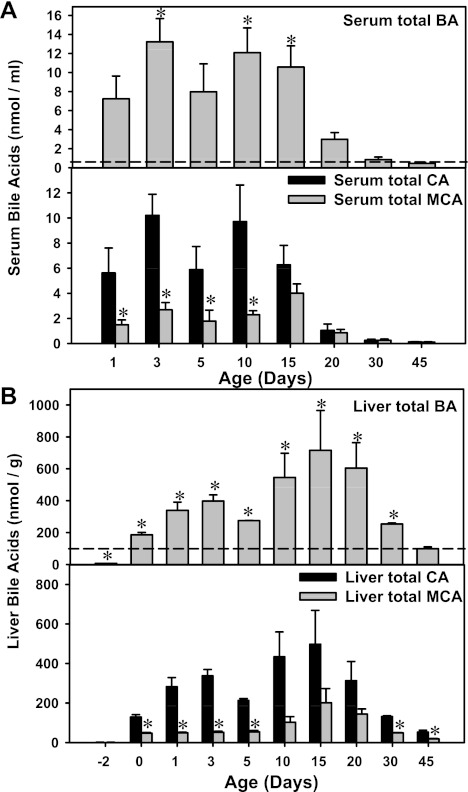
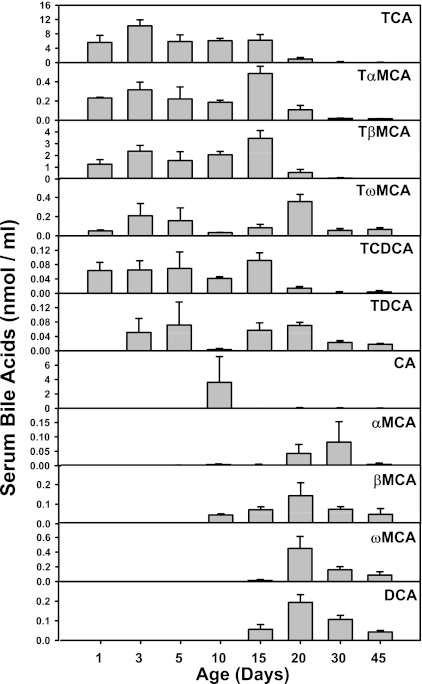
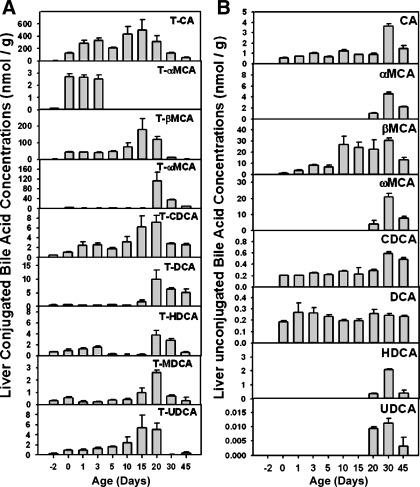
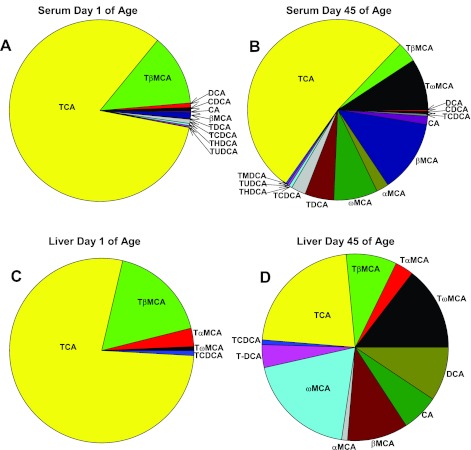
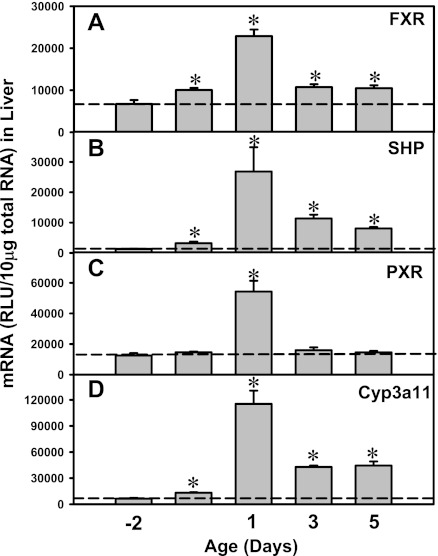
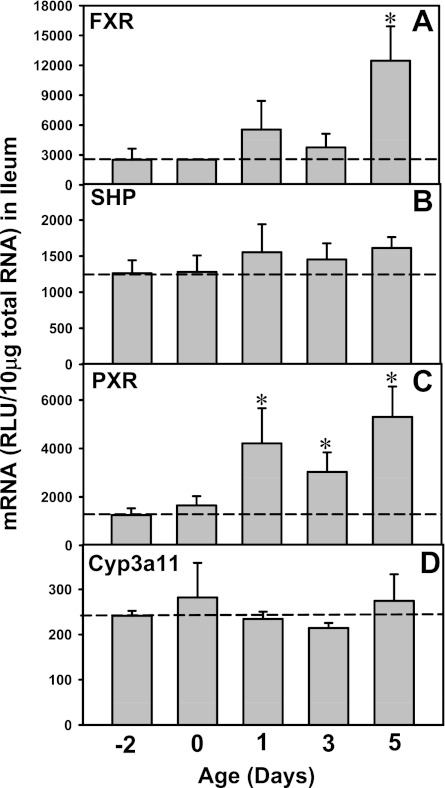
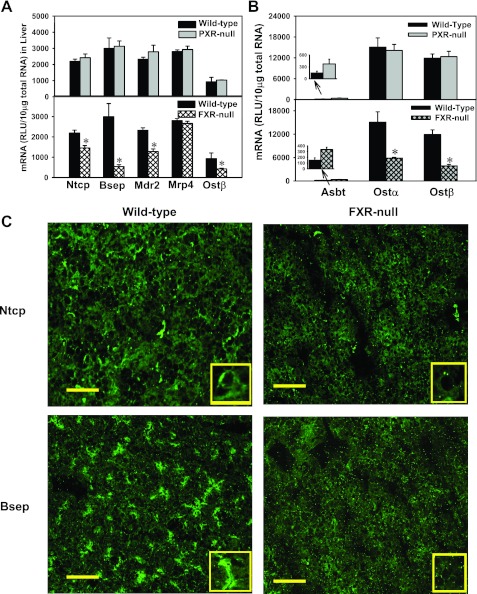
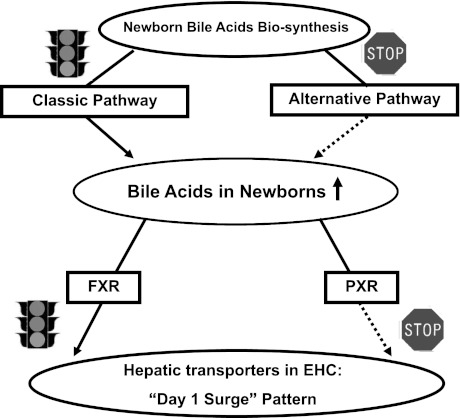
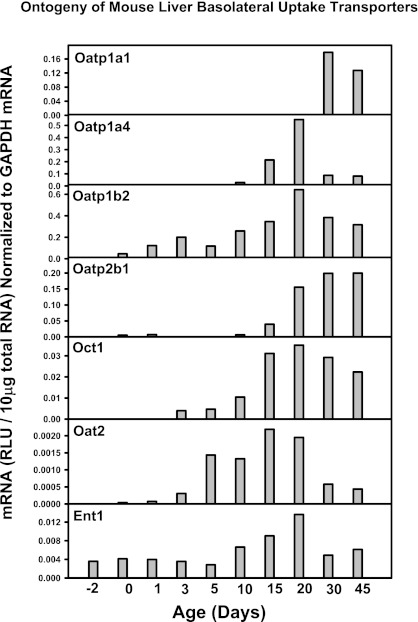
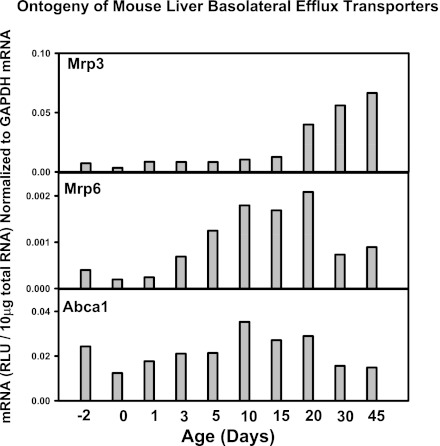
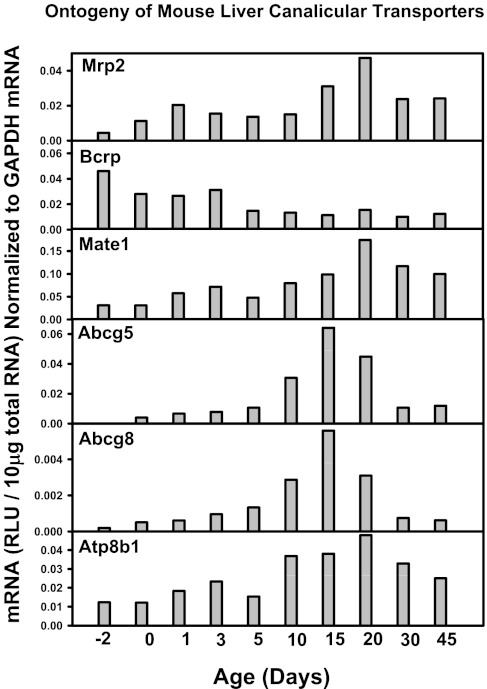
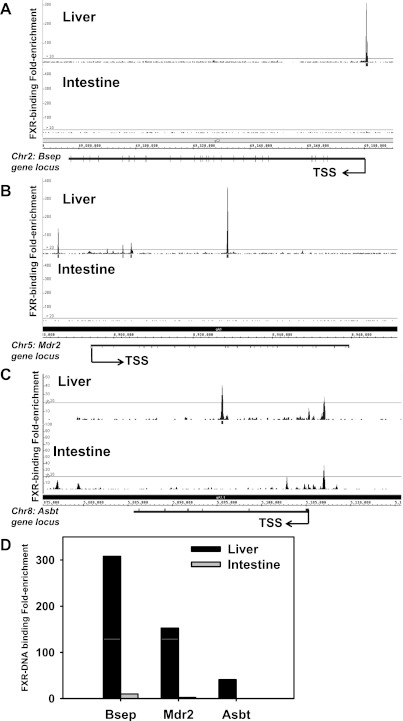
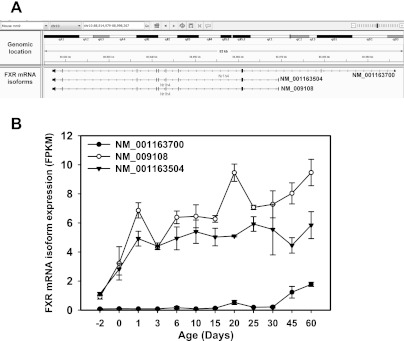
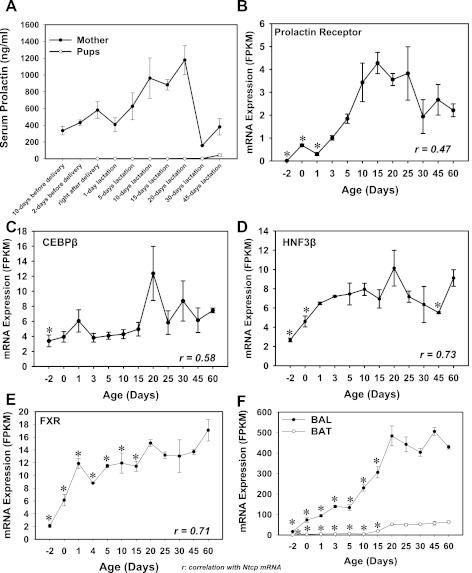
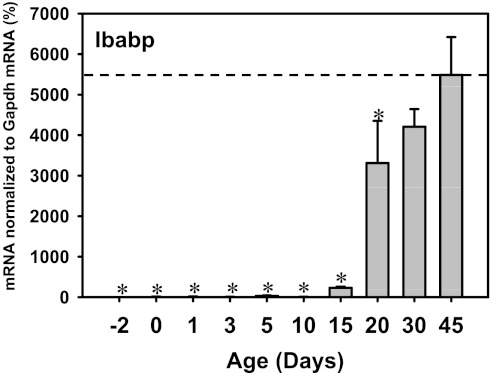
The enterohepatic circulation (EHC) of bile acids (BAs) plays a pivotal role in facilitating lipid absorption. Therefore, initiation of the EHC in newborns is of crucial importance for lipid absorption from milk. The purpose of this study was to determine at what age BA transporters in liver are expressed, and the mechanism for their initiation. Serum and liver samples were collected from C57BL/6 mice at 2 days before birth and various postnatal ages. Messenger RNA assays revealed a dramatic increase at birth in the expression of the BA transporters (Ntcp, Bsep, Mrp4, Ostβ), as well as the phospholipid floppase Mdr2 in mouse liver, with the highest expression at 1 day of age. The mRNA expression of the ileal BA transporters (Ostα and Ostβ) also markedly increased at birth. Meanwhile, taurine-conjugated cholic acid markedly increased in both serum and liver of newborns, correlated with upregulation of the classic pathway of BA biosynthesis in newborn liver. The mRNA levels of the major BA sensors, FXR and PXR, were increased at 1 day of age, and their prototypical target genes were upregulated in liver. The mRNA expression of transporters involved in the EHC of BAs was similar in wild-type and PXR-null mice. In contrast, in FXR-null mice, the "day 1 surge" pattern of Ntcp, Bsep, Ostβ, and Mdr2 was blocked in newborn mouse liver, and the induction of Ostα and Ostβ was also abolished in ileums of FXR-null mice. In conclusion, at birth, BAs from the classic pathway of synthesis trigger the induction of transporters involved in EHC of BAs in mice, through activation of the nuclear receptor FXR.
Figures
References
-
- Aleksunes LM, Slitt AM, Cherrington NJ, Thibodeau MS, Klaassen CD, Manautou JE. Differential expression of mouse hepatic transporter genes in response to acetaminophen and carbon tetrachloride. Toxicol Sci 83: 44–52, 2005 - PubMed
-
- Arrese M, Trauner M, Ananthanarayanan M, Boyer JL, Suchy FJ. Maternal cholestasis does not affect the ontogenic pattern of expression of the Na+/taurocholate cotransporting polypeptide (ntcp) in the fetal and neonatal rat liver. Hepatology 28: 789–795, 1998 - PubMed
-
- Assem M, Schuetz EG, Leggas M, Sun D, Yasuda K, Reid G, Zelcer N, Adachi M, Strom S, Evans RM, Moore DD, Borst P, Schuetz JD. Interactions between hepatic Mrp4 and Sult2a as revealed by the constitutive androstane receptor and Mrp4 knockout mice. J Biol Chem 279: 22250–22257, 2004 - PubMed
-
- Ballatori N, Christian WV, Lee JY, Dawson PA, Soroka CJ, Boyer JL, Madejczyk MS, Li N. OSTalpha-OSTbeta: a major basolateral bile acid and steroid transporter in human intestinal, renal, and biliary epithelia. Hepatology 42: 1270–1279, 2005 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases